Navsahyadri Institute of Pharmacy, (Naigaon) Nasarapur, Pune.
Insulin which is a polypeptide hormone that is secreted by pancreatic beta cells and is one of the important hormone for maintaining the blood glucose level. It also plays important role in maintaining the blood glucose level in the diabetic patients. Recent studies have showed that insulin also plays role in bone formation and attenuates in osteoporosis-related inflammation. Insulin structure consists of 51 amino acids, with chain A and chain B where these chains are connected by two disulphide bridges. Initiation of insulin synthesis takes place in beta cells of pancreas, with mRNA translation into preproinsulin having 110 amino acids, on transfer of this preproinsulin into endoplasmic reticulum it gets converted into proinsulin and after exiting from endoplasmic reticulum it moves into Golgi complex where it gets converted into mature insulin. Mature insulin is stored in the secretory granules until it combines with the plasma membrane to release the insulin. There are various types of inulin available in the market with different onset of and duration of action. This marketed synthetic insulin are called analogs of human insulin. Various guidelines are been proposed for storage and handling of the insulin. Also, the advancement in the recombinant DNA has made it possible for production of human insulin and insulin analogs by offering the improved efficiency, convenience and safety to the patients.
Insulin is a polypeptide hormone secreted by pancreas, by the beta cells present in islets of Langerhans [1]. In 1921-1922 Dr. Frederick Banting pioneered isolation and purification of the insulin in the Toronto. Insulin hormone plays important role in maintaing the blood glucose level, cell growth, and helps in the metabolism and is made of 51 amino acids [2]. Insulin is encoded by the INS gene in the humans, which is main anabolic hormone of body [4]. It regulates the uptake of glucose by the liver, skeletal muscles and the adipose tissues for the storage of excess glucose, where this absorbed glucose is then converted into the glycogen via the glycogenesis or fats via the lipogenesis [6]. As it an anabolic hormone, it coverts the small molecules present in blood to larger molecules in the cell. Low concentration of the insulin in blood has the opposite effect of the same, where the stored glucose is utilized from the various tissues and muscles. Production of insulin is high when there is greater amount of glucose present in blood as beta cells are sensitive towards the glucose, they secrete more insulin and inhibits insulin secretion when glucose levels are low [7]. Excess production and secretion of the glucose from the liver is controlled by the insulin, where the concentration of insulin in blood also helps with the synthesis of the protein in variety of the tissues. If pancreas don’t make insulin properly or the body does not utilize the insulin properly this can lead to the hyperglycemia. Which can result in type 2 diabetes [3]. As beta cells are sensitive to the glucose, they secrete insulin, where the neighboring alpha cells secrete glucagon that shows the opposite action as that of the insulin i.e. high concentration of glucagon when less glucose is present and low concentration when high glucose level is present [9]. The glucagon coordinates with the insulin to maintain blood glucose level, where insulin acts by the anabolic pathway and the glucagon acts by the catabolic pathway [1]. The glucagon helps to increase the blood glucose level by increasing the glycogenesis and gluconeogenesis in the liver [9,10]. So, the glucose homeostasis is maintained by regulating the secretion of insulin and glucagon from the alpha and beta cells in response to the blood glucose level [9]. Except diabetes recent surveys showed that insulin also acts on the other organs of the body and play different physiological roles like in brain, ear, kidney, bone, skin, and the hair follicles. Insulin also helps with the bone formation and also attenuates in the osteoporosis-related inflammation [8]. When there is absence of insulin or insulin activity this can lead to the diabetes, or the condition known as hyperglycemia, which causes the high blood glucose level. There are two types of diabetes depending on the insulin secretion and resistance. Beta cells are destroyed due to autoimmune response in the type 1 diabetes, so the body is not able to produce and secrete insulin [11]. Type 2 diabetes is caused primarily by the combination of the two factors like defective insulin secretion by the beta cells of pancreases and insulin resistance by the insulin sensitive tissues [12]. Insulin resistance can be said as clinical state where normal or elevated insulin level produces an attenuated biologic response [13]. Type 1 diabetes patients along with some type 2 diabetes patients requires the insulin therapy to achieve the optimal blood glucose level. As far the subcutaneous insulin therapy is the main stay of insulin therapy [5].
Structure Of Insulin:
Initially it was believed that the hormones would be the small molecules, until the structure of insulin was found to be quite large [2]. Human insulin consisting of the 51 amino acids, structure and the chemical property of insulin as polypeptide was found in 1928, where amino acid sequence of insulin was found in the year 1952. So, basically insulin is a dipeptide having the A and B chain which are linked by the disulphide bridges and having 51 animo acids. Insulin has the molecular weight of 5.8 kDa i.e. 941.176 g and its isotonic point is pH 5.5 [1]. Molecular formulae for human insulin is C257H383N65O77S6 [3]. So, in insulin Chain A is made up of 21 amino acids and chain B of 30 amino acids. There are two disulphide bridges (residues A7 To B7, and A20 to B19) that covalently connects the chains, and the chain A connects the internal disulphide bridge (residues A6 to A11). These joints are similar in all mammalian forms of the insulin. The strongly conserved sequence of insulin varies only slightly between species. Bovine insulin differs from human insulin only by three amino acid residues, and porcine by the one [4].

Figure 1: Structure of insulin
Biosynthesis Of Insulin:
Biosynthesis of insulin initiates with mRNA translation in the preproinsulin, which is a polypeptide having 110 amino acids with N terminal that is signal peptide, followed by the B chain then Connecting peptide (C peptide) and last C terminal as A chain (Fig. no. 2). On transfer of preproinsulin into the endoplasmic reticulum (ER) the signal peptide gets removed, and therefore converts the preproinsulin into the proinsulin, where the disulphide bond is formed between chain B and chain A. For the exit from ER the proinsulin moves through the Golgi complex towards the Trans Golgi network (TGN) to get divided into membrane enclosed organelles termed as secretory granules [1]. So, the division of C peptide in this compartment converts the proinsulin into the mature insulin having only Chain A and Chain B [2]. Mature insulin is stored in the secretory granules until it combines with the plasma membrane to release the insulin, or are degraded intracellularly by the autophagy or delivered directly to the lysosomes, other words describe as crinophagy [2,3]. Newly synthesized insulin is superiorly secreted [4,5]. Although multiple factors are involved in the biosynthesis of the insulin, metabolism of glucose is one of the important physiological events that stimulates the insulin gene transcription and m RNA translation [6].
Transcription and translation of insulin mRNA is regulated by the glucose. 20 folds increase in insulin levels is seen in glucose stimulated beta cells of pancreas [7,8].
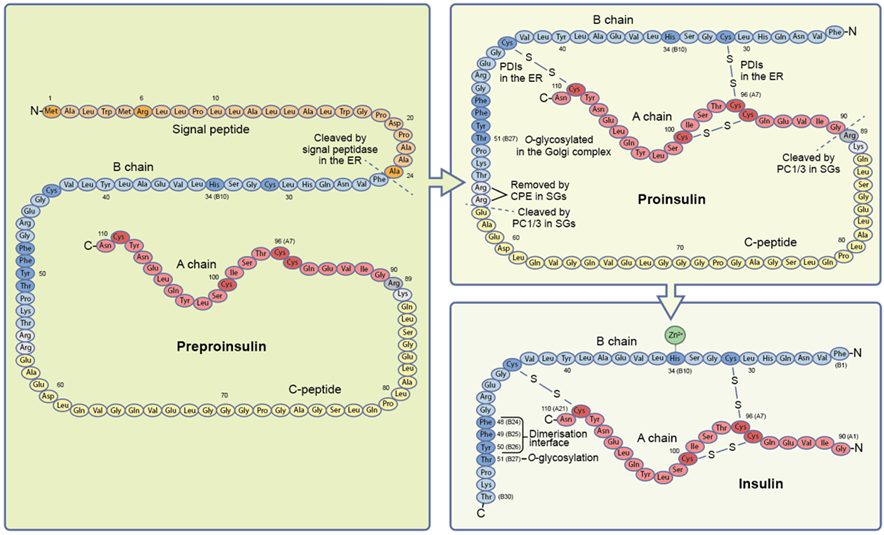
Figure 2: Biosynthesis of insulin
Regulation of preproinsulin mRNA stability:
The long half-life of preproinsulin mRNA is regulated by the conserved polypyrimidine tract and a UUGAAA- motif in its 3’ – UTR [9,11]. Preproinsulin mRNA stability is increased two to threefold by the rousing glucose levels, compared to the non-stimulated beta cells. Polypyrimidine tract binding protein 1 (PTBP1) is the best-known RNA binding protein that regulates the stability of the preproinsulin mRNA [12,13]. This PTBP1 binds to the 3’ – UTR of preproinsulin and avoids destabilization by opposing the T cell restricted intracellular antigen 1 related protein (TIAR) [14].
Glucagon like-protein 1 (GLP 1), which is released by nutrients stimulated L cells in gut, also helps in enhancing the stability of preproinsulin mRNA. Protein kinase A (PKA) mediated phosphorylation is induced when beta cells get exposed to GLP-1 [15], hnRNP K [12,14], hnRNP C, hnRNP E [12], hnRNP L, hnRNP U, HuD and poly(rC)- binding proteins (PCBP) 1,2 and 3 are the other preproinsulin RBPs present in the insulinoma cells [14]. Their involvement in the stability of preproinsulin mRNA is unknown [16]. The stability of preproinsulin mRNA is not expected to be affected in the type 2 diabetes, as the levels of preproinsulin mRNA in individual do not differ in type 2 diabetes and normoglycemia [17,18].
Regulation of preproinsulin mRNA translation:
Translation starts with binding of various initiation factors with 5/-UTR of mature mRNAs, in mammalian cells. This binding occurs in the specified manner and assists in placing large and small ribosomal subunits. On glucose stimulation the polysomes that stores preproinsulin mRNA in beta cells, are transported to ER, where the translation of preproinsulin starts immediately [19]. Along with the common regulators of translation, other specific factors regulate insulin translation with response to the nutrients, where ATP-dependent RNA helicase DEAD- box helicase 1(DDX-1) bind to the eukaryotic initiation factor (elF)3a and eIF4b and to preproinsulin mRNA [20]. These results can be useful in pathogenesis of type 2 diabetes, which is often linked with hyperlipidemia; saturated NEFAs, like palmitate increases the secretion of insulin [21]. And exposure of ER to palmitate causes the depletion in Ca2+ levels and causes the impairing in proinsulin folding and ER stress, further decreasing the insulin biosynthesis [22]. Translating ribosomes receives the amino acids from the transfer RNAs (tRNAs), and their post transcriptional methylation enhances the constancy and, efficiency of translation. Glucose intolerance due to impaired insulin synthesis can cause the mutation or polymorphism in the tRNA methyltransferases, like CDKAL1 and TRMT10A [23-25]. In case of TRMT10A it causes a young onset diabetes that is correlated with microencephaly and intellectual disability [25]. All eukaryotic mRNAs are capped at 5/-UTR and are translated in cap-dependent manner, sometimes cap-independent translation can occur due to placing of initiation factor closer to the first AUG codon by internal ribosome entry site (IRES). By avoiding control for cap-dependent translation, cap-independent translation permits the protein synthesis in conditions where the former is compromised, for example, upon conditions like hypoxia, irradiation, apoptosis or amino acid starvation. Cap-independent manner can be used for translating the mRNAs for insulin and other secretory granules, thus allowing their continuous production even in stress condition [15], and upon mTOR pathway inhibition [26]. Polypyrimidine tract-binding protein 1 (PTBP 1) is the key for this process, who’s binding to preproinsulin mRNA 5/-UTR increases the temporary hyperglycemia. Though longer exposure of human islets to hyperglycemia overpowers the PTBP 1 expression and insulin biosynthesis, this can be possibly due to upregulation of microRNA (miR)-133a that bind to 3/-UTR of the PTBP1 mRNA [27].
The hinderance in the folding at ER is due to the mutations in the insulin’s amino acid sequence, [28,29] that can be single point mutations or extensive exon deletion that can affects its translation. In such conditions the translation initiation of preproinsulin is seized, which leads to permanent neonatal diabetes. Translation initiation is also seized if there is mutation in the starting codon of preproinsulin. Impair ER translocation and target preproinsulin for proteasomal degradation also occurs if there is mutation in signal-peptide of preproinsulin, such as R6C replacement [30]. Mutations affecting signal-peptide cleavage, such as A24D replacement also causes permanent neonatal diabetes by blocking ER exit and causing ER stress [29,31].
Regulation of proinsulin folding:
Folding of proinsulin in ER is established by formation of three disulfide bonds, that are two interchain between A and B chain and one intrachain within A chain. Upon mutation of C96 in A chain caused due to alteration in cysteine paring can cause misfolding, accumulation and toxic aggregation of proinsulin in ER which can lead to permanent neonatal diabetes or mature-onset diabetes of the young (As seen earlier) [29].
In vitro studies have suggested that in human genome of among 15-20 protein disulfide isomerases (PDIs) found, the oxidative folding of proinsulin disulfide bonds on conjunction with the oxidoreductases ER oxidoreduction 1 α/β (ERO 1 α/β) is facilitated by PDIA 1 [31,32]. For successive rounds of disulfide bond generation, PDIs are regenerated by ERO 1 α/β. Even the healthy beat cells show moderate amount of proinsulin disulfide mispairing and in early type 2 diabetes the accumulation of misfolded proinsulin intermediates occurs [33,34]. Thus, for proper beta cell function maintaining the proper redox status which depends on continuous supply of reducing equivalents by cytosolic thioredoxin is important.
The antioxidative action of thioredoxin is inhibited by thioredoxin interacting protein (TXNIP) and this is elevated upon ER stress and insulin misfolding [35,36], elevation was also seen in type 2 islets [37]. Unfold protein response (UPR) counters the disturbance of proinsulin folding. From, the 3 UPR sensors, namely inositol-required enzyme 1 α (IRE1α), protein kinase RNA-like ER kinase (PERK) and activating transcription factor 6 (ATF6), IRE1α and PERK are involved in proinsulin folding [38]. IRE1α is active under certain physiological conditions for oxidative proinsulin folding [38], whereas PERK is induced by ER stress [39,40]. Wolcott-Rallison syndrome, an autosomal recessive permanent neonatal diabetes is caused due to mutation in PERK [39].
Regulation of proinsulin conversion to mature insulin:
Two steps are involved in conversion of proinsulin into mature insulin; first, cleavage of C- peptide junctions at B and A chain at basic residues R55-R56 and K88-K89 (Fig. no. 2) [41,42]. C-peptide is released from protein convertase 1/3 (PCI 1/3), in human beta cells [43]. Second, dibasic residues R55-R56 at C-terminal end of B chain is removed by exopeptidase carboxypeptidase E/H (CPE) (fig. no.2) [41,42]. While elevated proinsulin secretion along with impaired proinsulin conversion is the hallmark for type 2 [44] and type 1 diabetes [45].
Genetic variants that affect the junction between the C-peptide and A chain, such as replacement of R89 [29], or the proteolytic activities of either PC 1/3 or CPE [46] have also been found in few individuals with type 2 diabetes and altered glucose metabolism. Mature insulin is stored in secretory granules, in most of the mammalian beta cells including humans in the form of hexamer of three dimers each of which coordinates the binding of Zn2+ molecule to H34 (B10) in B chain (fig. no. 2). Although guinea pig insulin is exception to this as it does not bind to Zn2+. Zinc transporter 8 (ZT8) mediates the transport of Zn2+ in the secretory granules, where ZT8 is known autoantigen for type 1 diabetes and a risk gene for type 2 diabetes. Depletion in Zn2+ impairs insulin crystallization and insulin secretion in mice [47]. Proinsulin is converted to insulin by carriers of ZnT8 variants R325W that correlates with lower expression of transporter and have low chances of developing type 2 diabetes. Where in such individuals the import of Zn2+ in secretory granules is made by other zinc transporters [48,49].
Types Of Insulin Available:
There are different type of insulin available in market and each type works at different “onset” time, and its effects last for different time length, known as “duration. Peak is observed when the insulin shows the strongest effect. After peak, the effect of insulin wears off over the next few hours and so. Table below shows the type of insulin, how fast it starts to work (onset), when they reach peak level and how long it lasts (duration).
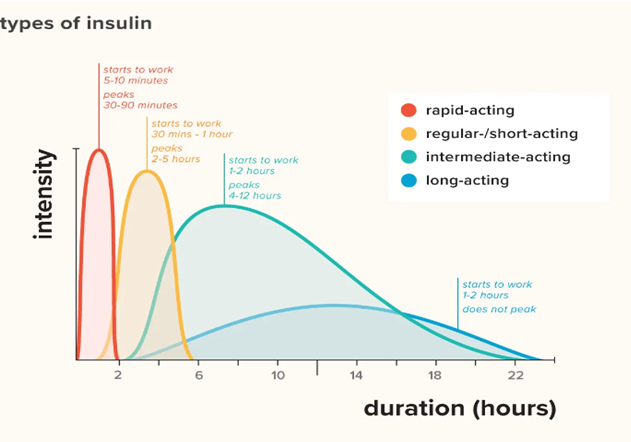
Figure.3: Types of insulin
Insulin analogs are the analogs that have been designed to mimic the body’s natural pattern of releasing insulin. This synthetic made insulin are called analogs of human insulin. However, the minor structural or amino acid changes in them give them the desirable characteristic when injected under the skin. Some of the examples of insulin analogs are mentioned below in the table along with their characteristic features.
Table No. 1: Types of insulin their brand name and duration of action
|
Category/Name of insulin |
Brand name (manufacturer) |
Alteration |
Preparations available |
Onset time |
Peak action |
Duration of action |
|
Short-Acting |
|
|
|
|
|
|
|
Regular Human |
Humulin R (Lilly) Novolin R (Novo Nordisk) |
- |
Vial
Vial |
~30 min. |
1.5-3.5 hrs. |
7-8 hrs. |
|
Rapid-Acting |
|
|
|
|
|
|
|
Insulin Lispro |
Humalog (Lilly) Admelog (Sanofi) Lyumjev (Lilly) |
Reversal of amino acid proline at B28 and lysine at B29. |
Vial, Cartridge, pen |
~15 min. |
30–70 min. |
2–5 hrs. |
|
Insulin Aspart |
Novolog, Novorapid, Flasp (Novo Nordisk) |
Replacing proline at B 28 with aspartic acid. |
Vial, Cartridge, pen |
10–20 min. |
1–3 hrs. |
3–5 hrs. |
|
Insulin Glulisine |
Apidra (Sanofi-Aventis) |
Replacing asparagine with lysine at B3 and glutamic acid with lysine at B29. |
Vial, pen |
10-29 min. |
~55 min. |
~6 hrs. |
|
Technosphere Insulin |
Afreeza |
- |
Inhaler |
- |
- |
- |
|
Intermediate-Acting |
|
|
|
|
|
|
|
NPH |
Humulin N (Lilly) Novolin R (Novo Nordisk) |
Neutral protaminated insulin. |
Vial, pen |
1.5-4 hrs. |
2.8-1.3 hrs. |
Up to 24 hrs. |
|
Long-Acting |
|
|
|
|
|
|
|
Insulin Detemir |
Levemir (Novo Nordisk) |
Myristic acid acylation to the lysine residue on position B-29 and Deletion of threonine from B30. |
Vial, pen |
1-2 hrs. |
6-8 hrs. |
Up to 24 hrs. |
|
Insulin Glargine |
Lantus (Sanofi-Aventis) Basaglar (Lilly) Toujeo (Sanofi-Aventis) |
Asparagine replaced with glycine at A21 and two arginine amino acids added at position B31 and B32 |
Vial, cartridge, pen |
1-3 hrs. |
No peak |
Up to 24 hrs. |
|
Insulin Deludec |
Tresiba (Novo Nordisk) |
Deletion of threonine at B30 and addition of 16?carbon fatty acid to lysine at B29 via a γ-L-glutamic acid linker. |
Pen |
0.5-1.5 hrs. |
No peak |
Up to 24 hrs. |
|
Insulin-Mixtures |
|
|
|
|
|
|
|
NPH/Regular Insulin 70/30 |
Humulin 70/30 (Lilly) Novolin 70/30 (Novo Nordisk) |
70% Isophane insulin and 30% Regular insulin. |
Vial, pen |
30-90 min. |
1.5-6.5 hrs. |
18-24 hrs. |
|
ProLispro/Lispro Humalog 75/25
|
Humalog Mix 75/25 (Lilly) |
75% neutral protaminated insulin lispro and 25% insulin lispro |
Vial, pen |
10-30 min. |
0.5-4 hrs. |
Up to 24 hrs. |
|
ProLispro/Lispro Humalog 50/50
|
Humalog Mix 50/50 (Lilly) |
50% neutral protaminated insulin lispro and 50% insulin lispro |
Vial, pen |
10-30 min. |
0.75-2 hrs. |
Up to 24 hrs. |
|
ProAspart/Aspart NovoMix 70/30 |
Novolog Mix 70/30 (Novo Nordisk) |
70% protaminated insulin aspart and 30% insulin aspart |
Vial, pen |
10-20 min. |
1-4 hrs. |
Up to 24 hrs. |
Guidelines For Insulin Storage And Handling:
Storage of insulin:
Handling of insulin:
There are different methods for insulin delivery such as syringes, injection pens, insulin pumps and inhalers.
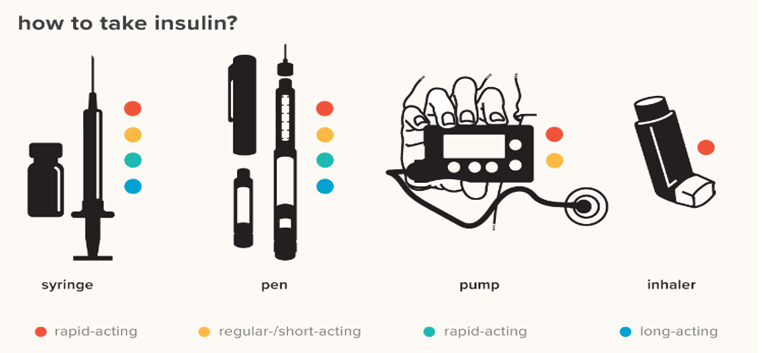
Figure 4: Methods for Insulin delivery
Role Of Insulin In Diabetes:
Insulin plays important role in treatment of diabetes either type 1 diabetes or type 2 diabetes. It helps to keep the blood glucose level in control and prevent the further complications related to diabetes. Insulin works similar to the human insulin hormone produced by the body. So, the role of insulin in human body is to control the blood sugar level and to store the extra glucose for the energy. If the person has diabetes, there blood sugar level keeps rising after they eat. That’s because their body don’t have enough insulin to store the glucose into the body cells. In type 1 diabetes the body stops producing insulin where in type 2 diabetes pancreas doesn’t make enough insulin, or in some patients the produced insulin is not utilized well by the body. Insulin therapy helps to keep the blood sugar level in the target range to avoid further complications like heart attack, stroke, kidney related diseases, eye problem, dental issues and nerve damage. In type 1 diabetes you need daily dose of insulin to stay healthy as your body doesn’t make enough insulin. In type 2 diabetes insulin can be the part of therapy in combination with other anti-diabetic drugs to manage the blood glucose level. Insulin therapy can also be needed in the type of diabetes that is developed during the pregnancy known as “gestational diabetes”.
CONCLUSION:
As insulin is a vital hormone in regulating the glucose metabolism, and its use in management of diabetes since its discovery has played an important role in many aspects. Understanding the biosynthesis and the structure of insulin has provided the various insulin formulation development for the treatment of diabetes according to the diverse patient’s needs. And effective diabetes management requires a comprehensive approach, incorporating insulin therapy, lifestyle modification and ongoing monitoring. Also, the advancement in the recombinant DNA has made it possible for production of human insulin and insulin analogs by offering the improved efficiency, convenience and safety to the patients.
REFRENCES




Sakshi Masaye*, Pooja Kashid, Trushali Mandhare, Kishore Otari, Lab to Life: The Journey of Insulin Synthesis and Its Types, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 3, 662-676. https://doi.org/10.5281/zenodo.14995654
 10.5281/zenodo.14995654
10.5281/zenodo.14995654
