*1Dept. of Pharmacology, School of Pharmaceutical Education and Research, Jamia Hamdard, New Delhi (110062), Delhi, India.
2,3,4Dept. of Pharmacology, School of Pharmaceutical Education and Research, Jamia Hamdard, New Delhi (110062), Delhi, India.
Thyroid hormones play a crucial role in regulating numerous physiological processes, yet their impact on reproductive and sexual health is often underestimated. Recent research suggests that imbalances in thyroid function-whether due to hypothyroidism or hyperthyroidism-can negatively affect the hypothalamic-pituitary-gonadal (HPG) axis, resulting in altered sex hormone levels, impaired gamete development, and reduced libido in both males and females. This article reviews recent clinical and experimental data to examine how thyroid imbalances contribute to infertility and sexual dysfunction, which affect millions worldwide. In men, thyroid dysfunction is frequently linked to lower testosterone levels, reduced sperm production, erectile difficulties, and issues with ejaculation. In women, thyroid hormone imbalances influence the development of ovarian follicles, menstrual cycle consistency, the receptivity of the endometrium, and pregnancy outcomes. Autoimmune thyroid conditions like Hashimoto’s thyroiditis and Graves’ disease add further complexity, particularly in women undergoing assisted reproductive treatments (ART). Fortunately, many of these reproductive and sexual symptoms can be reversed with timely correction of thyroid hormone levels, highlighting the importance of early diagnosis and management. In addition to their traditional hormonal roles, thyroid hormones act through both genomic and non-genomic pathways in reproductive tissues and interact with other hormonal signals such as leptin, prolactin, and insulin-like growth factors. These mechanisms offer potential pathways for developing novel diagnostic and treatment approaches. By integrating insights from both endocrinology and reproductive medicine, this review emphasizes the importance of evaluating thyroid function in individuals experiencing infertility or
The thyroid gland, shaped like a butterfly, is an endocrine structure situated beneath the thyroid cartilage in the neck of mammals [1]. It plays a central role in hormonal regulation as part of the hypothalamic-pituitary-thyroid (HPT) axis. As part of this hormonal regulatory system, the hypothalamus releases thyrotropin-releasing hormone (TRH), which in turn signals the anterior pituitary gland to produce and release thyroid-stimulating hormone (TSH). Thyroid-stimulating hormone (TSH) activates the thyroid gland to secrete the hormones thyroxine (T4) and triiodothyronine (T3), which exist in circulation either freely (biologically active) or bound to proteins (inactive) [2] (Figure 1). Within the bloodstream, T4 is mainly transformed into the more potent T3 form through enzymatic action by type I and type II deiodinases, predominantly located in tissues such as the liver, kidneys, muscles, and thyroid [3,4]. Approximately 80% of the T3 found in circulation originates from the conversion of T4 via the process of deiodination [5]. These thyroid hormones (THs) then travel swiftly to various tissues, where they regulate energy metabolism and support essential physiological processes, including those of the reproductive system.
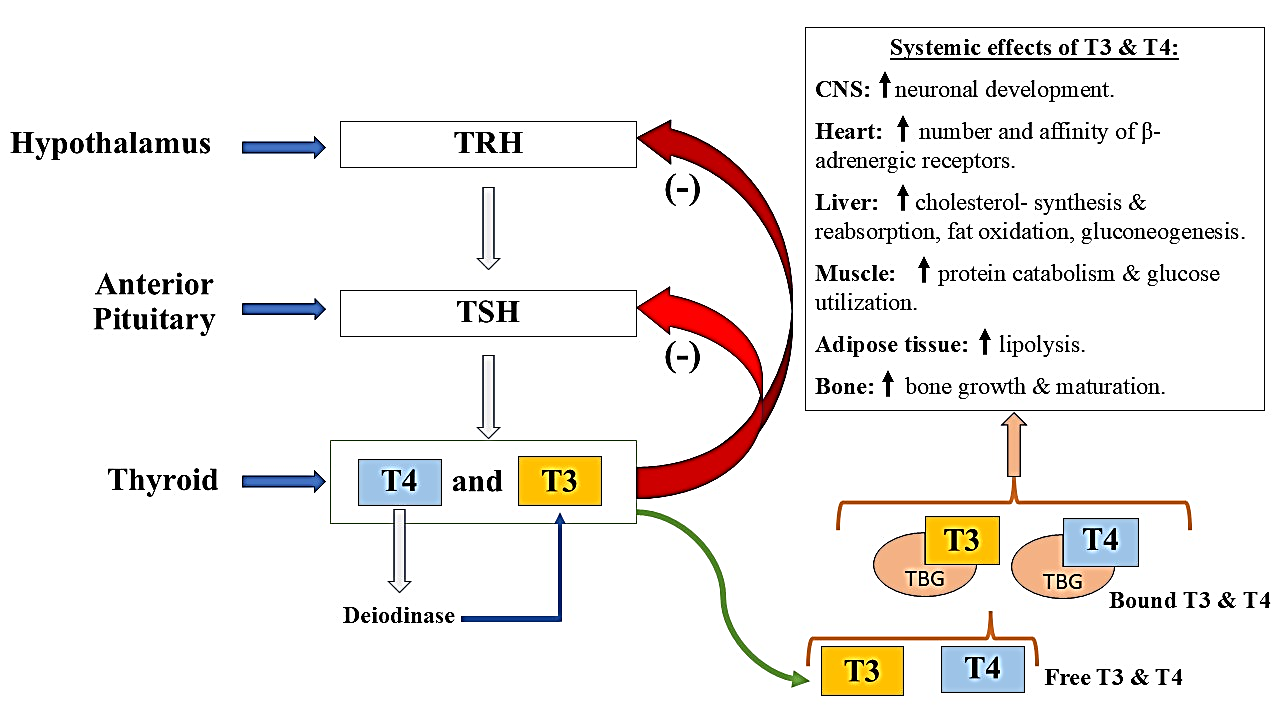
Figure 1: Illustration outlining the functional hypothalamic-pituitary-thyroid (HPT) axis, including the physiological effects of circulating and protein-bound forms of tri-iodothyronine (T3) and thyroxine (T4) throughout the body. Abbreviations: TBG– thyroxine-binding globulin; TRH– thyrotropin-releasing hormone; TSH– thyroid-stimulating hormone.
Thyroid disorders such as hypothyroidism and hyperthyroidism are frequently encountered in clinical practice and manifest through a wide range of symptoms that are well documented. The vast majority-about 95%-of hypothyroidism cases are due to dysfunction within the thyroid gland itself, and these are typically categorized as either overt or subclinical forms. In regions where iodine consumption is sufficient, the leading cause of hypothyroidism is autoimmune thyroiditis, commonly known as Hashimoto’s thyroiditis [6]. On the other hand, hyperthyroidism arises from a broader range of causes, with Graves’ disease being the most prevalent. This autoimmune disorder results from antibodies that imitate TSH, leading to excessive stimulation of the thyroid gland [6].
Data from the U.S. National Health and Nutrition Examination Survey III report that 4.6% of individuals are affected by hypothyroidism, with 0.3% presenting with overt disease and 4.3% with subclinical hypothyroidism. Women are significantly more likely to be affected, showing prevalence rates 5 to 8 times greater than those seen in men [7]. Women are disproportionately affected, with rates 5-8 times higher than in men [7]. While overt hypothyroidism in men is rare (0.1%), subclinical hypothyroidism still has a notable prevalence of 2.8% [8]. The same national survey reported that hyperthyroidism affects approximately 1.3% of the population, with overt cases comprising 0.5% and subclinical forms 0.7%. As with hypothyroidism, women are more frequently affected than men [7].
As outlined in Table 1, the symptom profiles of both thyroid conditions are well characterized. Diagnosis typically involves measuring serum TSH alongside free T4 and/or T3. A diagnosis of hypothyroidism is confirmed by elevated TSH levels (>5.0 mU/L) and low levels of free T4 or T3 [9], while hyperthyroidism presents with low TSH (<0.4 mU/L) and elevated free thyroid hormone levels [10].
Table 1: Common clinical signs and symptoms noted in patients with hypothyroidism or hyperthyroidism [11,12].
|
Underactive Thyroid (Hypothyroidism) |
Overactive Thyroid (Hyperthyroidism) |
|
Constant fatigue and trouble concentrating |
Feelings of agitation and mood instability |
|
Discomfort in cold weather |
Difficulty tolerating heat |
|
Weight gain without obvious cause |
Weight loss accompanied by a reduced desire to eat |
|
Skin tends to be dry and cool to the touch |
Warm, moist skin is common |
|
Slowed heartbeat |
Accelerated heart rate and palpitations |
|
Menstrual periods that are heavy or last longer than usual |
Irregular menstruation or absent periods |
|
Muscle fatigue, stiffness, or numbness |
Muscle tremors and general weakness |
|
Constipation or infrequent bowel activity |
Increased frequency of bowel movements or diarrhoea |
|
— |
Swelling of the thyroid and protruding eyes |
Subclinical thyroid dysfunction, characterized by abnormal TSH levels with normal T4 and T3, can still exert significant physiological effects [10]. As elaborated later in this review, such dysfunctions may adversely affect sexual health in both men and women.
Standard treatment for hypothyroidism involves levothyroxine (synthetic T4) replacement therapy, which usually requires 4-6 weeks to normalize hormone levels. For hyperthyroidism, treatment options include radioactive iodine therapy, antithyroid drugs, or surgical removal of the thyroid gland.
1.1 Thyroid hormones production, transport, and mechanisms of action:
Thyroid hormones (THs) are synthesized in the thyroid gland and circulate through the bloodstream; however, their tissue-specific availability is predominantly regulated by a group of enzymes known as deiodinases [13]. There are three primary forms of these enzymes: DIO1, DIO2, and DIO3. DIO2 is responsible for converting the prohormone thyroxine (T4) into the active hormone triiodothyronine (T3), while DIO3 deactivates both T3 and T4. DIO1 has a dual role-it can either activate or deactivate THs-but it exhibits lower specificity and requires high substrate concentrations to convert T4 into T3 effectively [14,15]. Although T3 and T4 are lipid-soluble, they require the assistance of specific transport proteins to enter cells. These transporters include members of the monocarboxylate transporter (MCT) family-namely MCT8/SLC16A2 and MCT10/SLC16A10-as well as organic anion transporting polypeptides (OATPs), such as SLCO1C1 and OATP1C1 [16]. In murine models, Mct8 appears more essential than Mct10, as Mct8-deficient mice exhibit altered thyroid hormone profiles and tissue-specific imbalances, unlike Mct10-deficient mice [17]. In humans, mutations in the MCT8 gene result in Allan-Herndon-Dudley syndrome, a condition characterized by profound neurological deficits [18]. Interestingly, mice lacking Mct8 do not show such severe manifestations, possibly because they can still utilize T4 via Oatp1c1, which is then locally converted to T3 [17].
The actions of thyroid hormones are primarily mediated through their interaction with nuclear thyroid hormone receptors (THRs), specifically THRα and THRβ. These receptors remain inactive until bound by THs. Upon activation, THRs bind to specific DNA sequences known as thyroid hormone response elements (TREs), typically as heterodimers with retinoid X receptors (RXRs) [19]. TREs comprise two repeated sequences (AGGT/ACA), arranged in different configurations such as direct repeats or palindromes, where RXR generally binds the 5′ site and THR the 3′ site [20]. In the absence of hormone, THRs recruit corepressors to inhibit gene transcription. Hormone binding causes a conformational change in THRs, releasing corepressors and attracting coactivators, thereby enabling gene transcription [21,22] (Figure 2). The typical configuration of DNA binding sites includes a four-base pair spacer (DR4), although variations exist. Triiodothyronine (T3) exhibits nearly tenfold greater binding affinity for thyroid hormone receptors (THRs) compared to thyroxine (T4) [21]. Beyond these genomic functions, THs also exhibit non-genomic effects, meaning they influence cellular functions without directly affecting gene transcription. These actions occur rapidly-within seconds to minutes-compared to genomic responses [26]. Initial studies showed that T3 could associate with membranes of rat erythrocytes and liver mitochondria [27]. Further investigations demonstrated that THs can modulate ATP generation, oxygen consumption, pH levels, and ion transport [28]. These rapid effects contribute to maintaining cellular homeostasis, including ionic balance and structural integrity. Evidence also suggests a dynamic interplay between genomic and non-genomic actions, indicating a complex regulatory network [23]. Although the precise molecular mechanisms of non-genomic effects remain to be fully elucidated, these effects are initiated at the plasma membrane or in the cytoplasm [25]. Some involve a membrane-associated form of THRα that targets specific membrane regions after undergoing palmitoylation.
Moreover, THs may also act through the integrin αVβ3 receptor located on the cell surface. Stimulation of this receptor can enhance the expression of genes like FGF2, HIF1α, and THRA, while suppressing genes such as CASP3 and APAF1 [24]. These responses are thought to be more strongly elicited by T4, although it remains unclear whether T3 or T4 is the primary ligand for this receptor [24]. While direct evidence for sex-specific non-genomic TH effects is lacking, the fact that THs modulate immune responses-which are themselves sex-dependent-implies potential sex-related differences [29]. For instance, THs are critical for brain development, supporting processes such as neuronal growth and cytoskeletal dynamics through non-genomic pathways [30]. T3 also appears to promote neuroprotection by activating signaling molecules like Akt and nitric oxide synthase. Future studies are necessary to understand how these pathways may contribute to sex-based variations in brain function. The regulation and production of thyroid hormones are primarily governed by the hypothalamic-pituitary-thyroid (HPT) axis. The hypothalamus, specifically the paraventricular nucleus, secretes thyrotropin-releasing hormone (TRH), which is delivered to the anterior pituitary via the portal circulation. There, TRH induces the release of thyroid-stimulating hormone (TSH). Thyroid-stimulating hormone (TSH) acts on the thyroid gland, prompting it to synthesize and release thyroid hormones. These hormones, in turn, regulate TRH and TSH secretion through a negative feedback mechanism to maintain hormonal equilibrium [31].
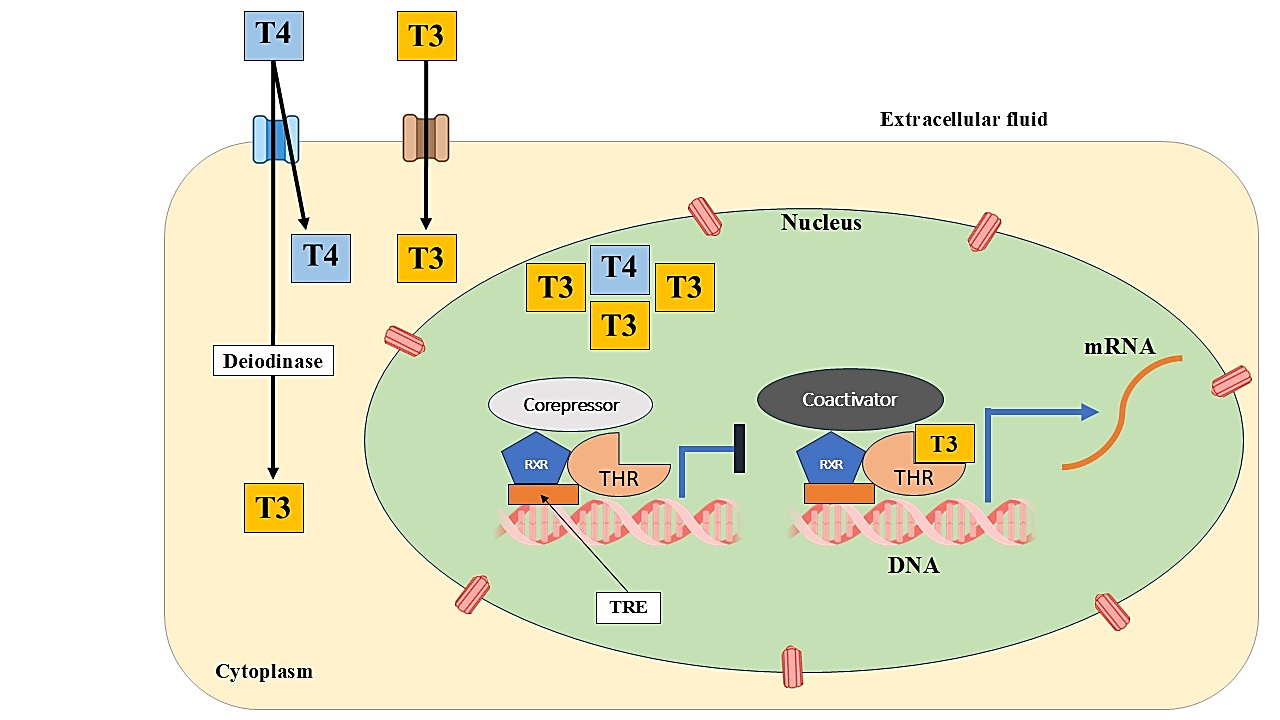
Figure 2: Depiction of the intracellular signaling mechanism of thyroid hormones (TH). Cellular uptake of TH is facilitated by specific transport proteins. Once inside the cell, T4 may be converted to T3 by the enzyme DIO2. Although not depicted, DIO1 also plays a role in this conversion. T3 and T4 interact with thyroid hormone receptors (THRs), which are primarily bound to retinoid X receptors (RXRs). This binding induces a structural change in the THRs, resulting in the release of corepressors and the recruitment of coactivators, thereby initiating the transcription of genes located downstream of thyroid hormone response elements (TREs).
1.2 Overview of sexual dysfunction in males and females:
Significant disturbances in the sexual health of both men and women have been linked to thyroid disease. A complex and common disorder that affects both sexes, sexual dysfunction can have several underlying causes. It typically comprises of several connected symptoms, sometimes including several different illnesses. It may be difficult to tell certain symptoms or diseases apart because they might manifest in similar ways [32]. The International Index of Erectile Function (IIEF), which was first introduced in 1997, weighs five main factors to measure male sexual health: erectile function, orgasm, sexual desire, contentment with intercourse, and overall sexual satisfaction. Despite limitations, this technique is highly validated across cultures and is regarded the baseline for measuring male sexual function (Figure 3) [33–37].
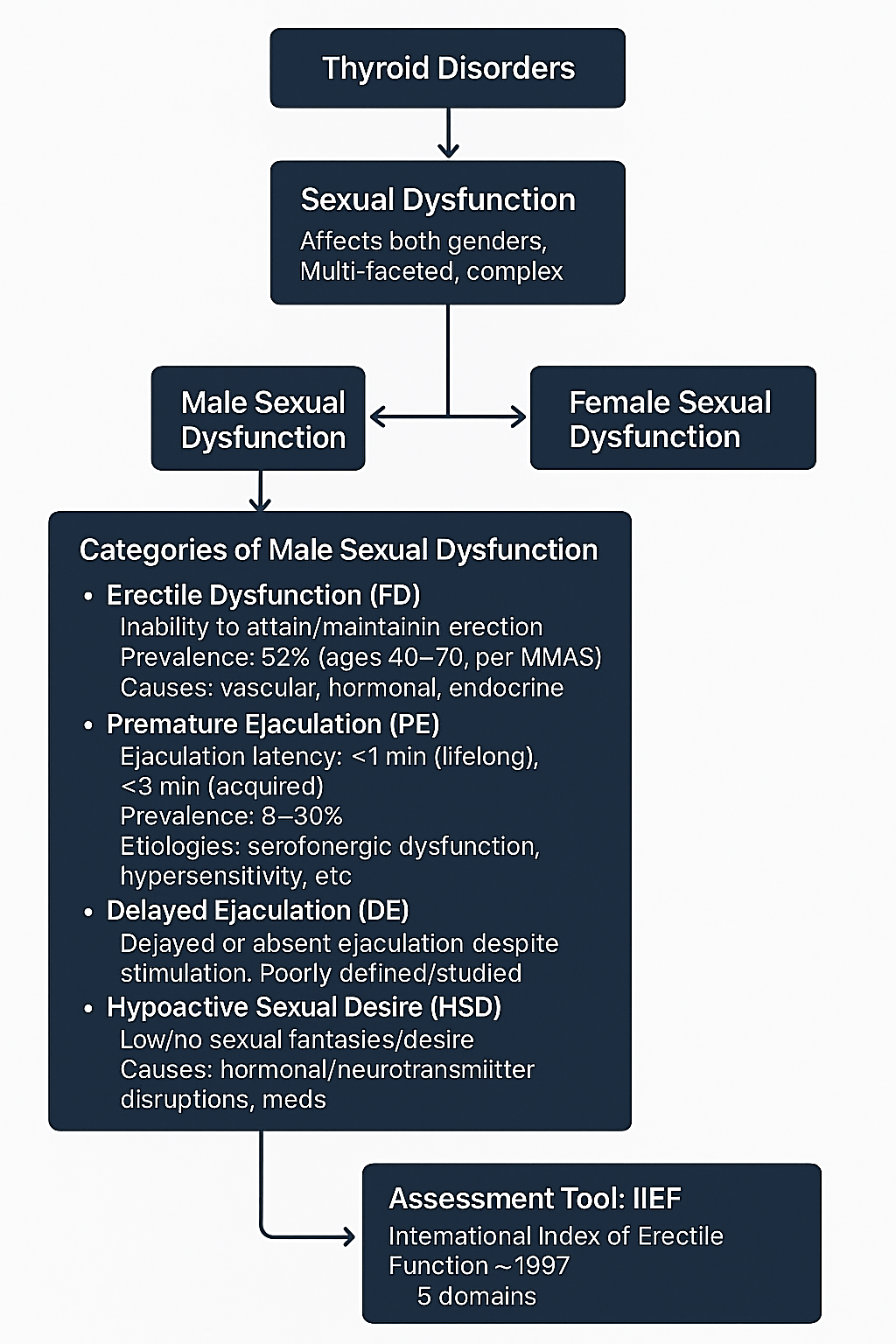
Figure 3: Overview of how thyroid hormone imbalances influence male sexual function.
The Figure 4 shows how sexual desire, arousal/lubrication, orgasm, overall satisfaction, and pain experienced during sexual intercourse are the criteria used to classify female sexual dysfunction [35,38].
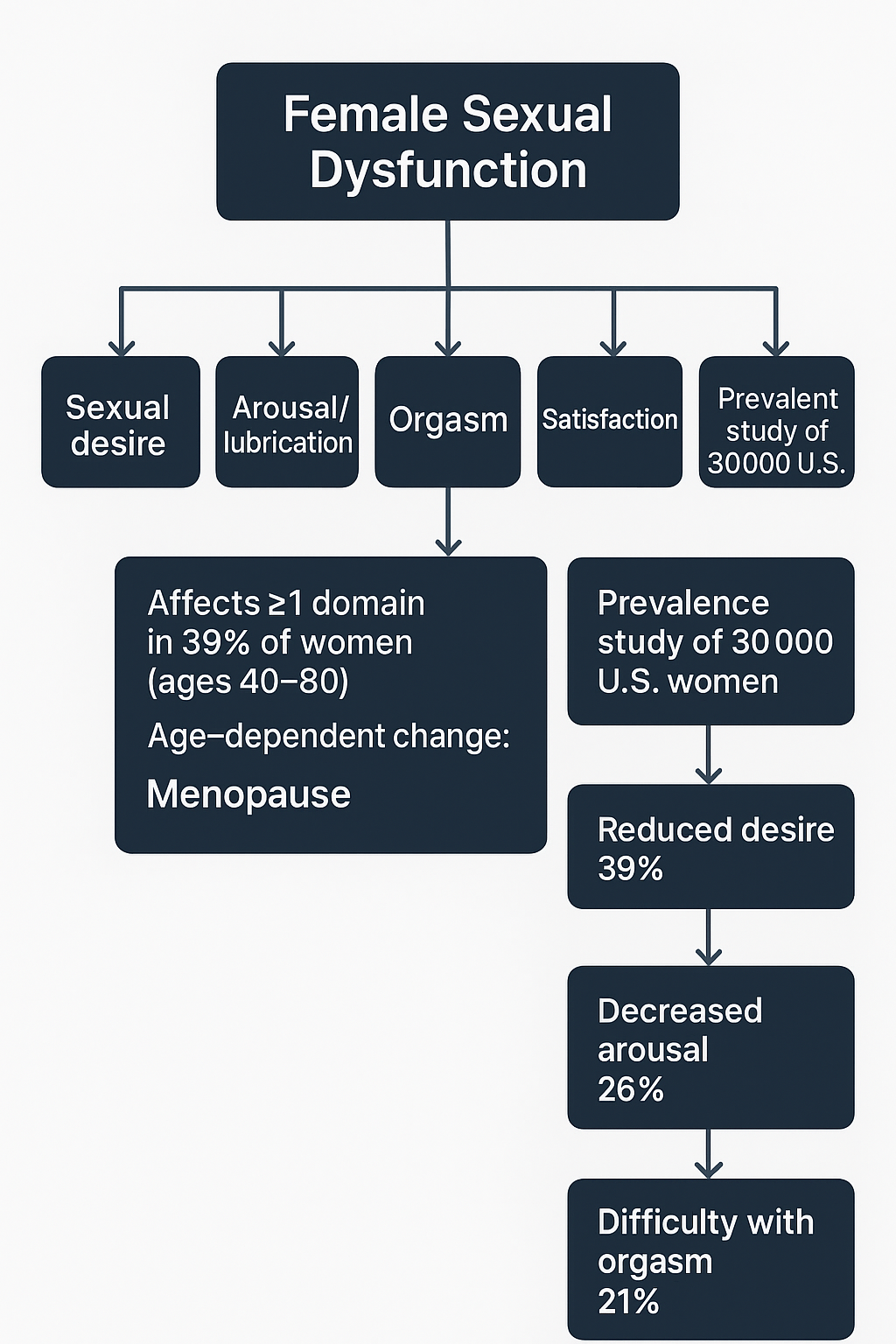
Figure 4: Overview of how thyroid hormone imbalances affect female sexual function.
Numerous environmental and lifestyle factors interact intricately to cause internal physiological changes that result in male infertility or subfertility [39]. The thyroid hormone profile, which serves a variety of biological functions [40] and is essential for preserving a healthy male reproductive capacity, is one of the major variables affecting reproductive health. Semen parameters and reproductive potential have been demonstrated to be adversely affected by changes in thyroid hormone levels [41]. Thyroid hormones' function in controlling male reproduction has been the subject of numerous studies [42], although little is known about the precise mechanisms by which thyroid abnormalities affect fertility indicators [43].
Transcriptional regulatory mechanisms are the main way that thyroid hormones affect the body. Sertoli cells in both foetuses and adults have been found to contain nuclear receptors for these hormones [44]. Activating gene expression and protein production, triiodothyronine (T3) stimulates Sertoli cell proliferation and differentiation when it attaches to its receptors within testicular cells [45]. Conversely, thyroid hormones can have non-genomic effects by interacting with receptors outside the nucleus, which may change the structure, metabolism, and proliferation of cells. There is still much to learn about the precise differences between genomic and non-genomic pathways [46]. Research indicates that cyclic adenosine monophosphate (cAMP) synthesis, calcium release, and sperm motility can all be improved by non-nuclear binding of thyroid hormones. Significantly, supplementing semen samples with thyroxine (T4) resulted in increased sperm hypermotility within 20 minutes [47]. Furthermore, by interacting with the mitochondria or cytoskeletal components of sperm cells, iodinated thyroid chemicals may have unknown non-genomic effects on them [46]. Thyroid hormones help maintain antioxidant defences and regulate oxidative equilibrium [48]. Neonatal hypothyroidism, for example, has been associated with increased oxidative stress, which interferes with the control of glucose in the testes and ultimately affects the survival, proliferation, and expression of proliferating cell nuclear antigen in germ cells [49]. Protein carbonyls and other indicators of oxidative stress rise in transitory hypothyroidism, although antioxidants such as catalase, glutathione reductase, glutathione peroxidase, and superoxide dismutase (SOD) fall. Only catalase and SOD levels, however, seem to increase in chronic hypothyroidism [50].
Reduced blood testosterone concentrations have been demonstrated to be positively correlated with lower thyroid hormone levels [51]. Erectile dysfunction, delayed ejaculation, decreased sexual drive, and poor semen quality are some of the male reproductive issues associated with this hormonal imbalance. The exact processes by which hypothyroidism influences semen qualities are still unclear, despite the fact that numerous studies have linked it to male infertility [51].
The fundamental process entails a drop in circulating T3 and T4 levels, which lowers testosterone and SHBG and ultimately interferes with spermatogenesis. Sexual dysfunctions like erectile dysfunction, delayed ejaculation, decreased libido, and poor semen quality can result from a prolonged thyroid hormone deficiency during childhood or puberty that suppresses the pituitary's release of gonadotropin, which in turn affects testicular development, function, and accessory gland secretions. Furthermore, it results in decreased testicular, epididymal, and prostate weights as well as lower seminiferous tubule widths, all of which hinder sperm production, motility, and passage through the male reproductive system. Oxidative stress brought on by hypothyroidism may further reduce sperm viability [58].
Excessive blood levels of sex hormone-binding globulin (SHBG) and a slower metabolic clearance of serum testosterone are associated with hyperthyroidism, which is characterized by excessive serum total T4 levels, according to human studies [59]. Men with hyperthyroidism frequently exhibit higher amounts of circulating estrogen and lower levels of bioavailable testosterone. Symptoms including erectile dysfunction, decreased sexual desire, and gynecomastia can be exacerbated by these hormonal abnormalities. Furthermore, men with hyperthyroidism exhibit heightened luteinizing hormone (LH) and follicle-stimulating hormone (FSH) responses to external gonadotropin-releasing hormone (GnRH) treatment. Testicular function is subsequently compromised, and semen quality is impacted by these hormonal changes.
Despite these results, hyperthyroidism and semen quality have only been directly associated in a small number of clinical studies. For instance, two of the three hyperthyroid male participants in a research by Clyde et al. exhibited oligozoospermia along with decreased sperm motility, and the third individual had both low sperm count and impaired motility [62]. Similarly, Kidd et al. found that hyperthyroid individuals had sperm concentrations less than 40×10?/mL [60]. A 1992 research by Hudson and Edwards with 16 hyperthyroid men reported similar results [63]. 9 of the 21 hyperthyroid males in a different study by Abalovich et al. had lower sperm counts, 18 had poor sperm motility, and 13 had abnormalities in progressive motility [61]. Krassas et al. assessed semen parameters in 23 hyperthyroid men and 15 euthyroid controls in a prospective research. The individuals' semen was examined both before and after receiving methimazole treatment, either by alone or in combination with radioiodine. Comparing the results to the control group, there were slight decreases in sperm density, alterations in sperm morphology, and a notable drop in motility. Crucially, following therapy, sperm motility and concentration increased [64]. The conditions asthenozoospermia, oligozoospermia, and teratozoospermia have all been closely linked to thyrotoxicosis, which results in high amounts of thyroid hormones in the blood. Hypoposia, or decreased semen volume, may also be a result of elevated thyroid hormone levels [61].
On the other hand, little is known about the relationship between hyperthyroidism and semen quality in animal models. Nonetheless, the research that is now available indicates that hyperthyroid rats might have smaller seminiferous tubule widths, decreased mitochondrial efficiency, and a delayed initiation of spermatogenesis. According to one study, hyperthyroidism in mice raised catalase activity while lowering glutathione peroxidase activity [57].
2.2 Thyroid disorders and reproductive dysfunctions in males:
Thyroid dysfunction affects the production of androgen-binding protein (ABP) in Sertoli cells and sex hormone-binding globulin (SHBG) in the liver, both of which are involved in the transportation of testosterone. Increased levels of SHBG and testosterone are seen in thyrotoxicosis patients, most likely as a result of decreased metabolic clearance. Nevertheless, in vitro research revealed that Sertoli cells produced less ABP when exogenous T3 was added [65].
Ejaculation is regulated in part by thyroid hormones and other metabolic regulators. Hypothyroidism has been associated with delayed ejaculation, but hyperthyroidism is more commonly associated with premature ejaculation. Male hypothyroids had greater rates of decreased libido, erectile dysfunction, and delayed ejaculation (64.3%), according to a multi-center study assessing sexual dysfunction in people with thyroid diseases. In contrast, only 7.1% of men with hypothyroidism reported premature ejaculation, whereas 50.0% of men with hyperthyroidism did. About half of the hypothyroid respondents experienced an improvement in delayed ejaculation, while the incidence of premature ejaculation decreased to 15% once thyroid hormone levels were stabilized in hyperthyroid patients [66]. An additional survey that used the Sexual Health Inventory for Males (SHIM) revealed a significant disparity in sexual function scores between thyroid dysfunction patients and healthy controls: 78.9% of those with thyroid dysfunction scored 21 or lower, which is indicative of dysfunction, while only 33.8% of the control group did the same. Both hyperthyroid and hypothyroid males showed significant improvements in their SHIM scores after treatment (P<0.0001). Furthermore, compared to control groups, a significantly greater prevalence of erectile dysfunction was noted in people with thyroid problems [56].
Thyroid hormones may have a direct impact on spermatogenesis and are essential for testicular growth and function. Thyroid hormone excess or insufficiency, as well as variations in thyroid levels, may have a variety of effects on male reproductive physiology, according to clinical and experimental research, especially in male rats and humans. For instance, larger testes have been linked to temporary hypothyroidism in rats during the neonatal stage [67]. When compared to euthyroid controls, hyperthyroid men have clinically demonstrated decreased sperm motility and conditions including asthenozoospermia, oligozoospermia, and teratozoospermia. Higher free T3 levels have recently been associated with higher serum fructose concentrations and ejaculate volume [68]. It has been demonstrated that hypothyroidism affects sperm motility and morphology, which raises teratozoospermia indices. Sperm count, motility, morphology, and seminal vesicle size were all considerably impacted in hypothyroid individuals. In mouse models of hypothyroidism, there was a drop in testicular germ cell populations, a reduction in sperm production, and a decrease in viable epididymal spermatozoa [57]. Other effects include decreased sperm motility and fertilization potential, impaired secretory function of the epididymis, and decreased expression of androgen receptors in the gonads. Reduced acrosome integrity, altered lipid peroxidation, mitochondrial apoptosis, and general mitochondrial activity are all indicators of compromised mitochondrial function [50,57]. The crucial metabolic involvement of thyroid hormones in sperm generation is shown by hypothyroidism's increased expression of thyroid hormone receptor alpha-1 (Thra1) and decreased expression of the deiodinase enzyme Dio3 [57]. Conversely, hyperthyroid rat models have been found to exhibit impaired mitochondrial activity and oxidative balance, delayed spermatogenesis, inhibited maturation, Leydig cell hyperplasia, and narrowing of seminiferous tubules. Additionally, elevated expression of solute carrier family 16 member 2 and monocarboxylate transporter 8 (MCT8), both essential for plasma membrane integrity, was linked to hyperthyroid conditions [56].
2.3 Thyroid disorders and testicular oxidative stress:
It appears that hyperthyroidism makes the tissues more susceptible to oxidative injury. Given that thyroid hormones frequently activate physiological processes, hyperthyroidism is bound to overstimulate metabolic states and increase the production of excess free radicals, which can lead to oxidative damage and lipid peroxidation in various tissues [69]. In comparison to other tissues, the testis is far more vulnerable to oxidative damage due to its high content of unsaturated fatty acids, strong machinery that produces reactive oxygen species, and low antioxidant capacity. According to studies, different thyroid hormone isomers may cause variable amounts of oxidative stress when hyperthyroidism is induced. As evidenced by increasing amounts of malondialdehyde, thiobarbituric acid reactive compounds, lipid hydroperoxide, protein carbonyl contents, or hydrogen peroxide, it has been proposed that hyperthyroidism causes a significant rise in testicular oxidative stress [69]. When hypothyroid patients received short-term L-thyroxine treatment, changes in antioxidant defense measures, such as elevated testicular glutathione (GSH) concentrations, were noted. Following three days of T3 administration, oxidized glutathione disulfide levels rose and GSH levels fell, resulting in a drop in the ratio of reduced to oxidized glutathione in the testis. Testicular GSH levels in both mitochondrial and post-mitochondrial fractions increased after five days of T3 administration, as did testicular ascorbic acid levels [69]. The "reduced to oxidized" glutathione ratio was elevated in both the mitochondrial and post-mitochondrial portions of hyperthyroidism caused by L-thyroxine or T3 [50,69]. There have been reports linking acute hyperthyroidism to elevated testicular catalase (CAT), glutathione peroxidase (GPx), and glucose-6-phosphate dehydrogenase activity and decreased testicular SOD [50,69]. L-thyroxine has been demonstrated to cause a multifold rise in GPx in the testis' mitochondrial and post-mitochondrial fractions [70]. T3 treatment increased seleno-independent GPx activity only in the testicular mitochondrial fraction [70]. These increased seleno-dependent and/or independent GPx levels, which are reactions to hyperthyroidism induced by L-thyroxine [70] or T3 [69], could be viewed as an adaptive reaction to counteract the harmful effects of hydrogen peroxide on the testicles. Male fertility is impacted by changes in oxidative stress parameters and testicular antioxidant defense systems brought on by hyperthyroidism, which result in a decrease in the number of viable and total sperm [50,69].
Likewise, because of a depressed metabolic state, hypothyroidism impairs antioxidant capacity, raises the production of reactive molecules, and upsets testicular redox equilibrium. Variations in the antioxidant defense system during testicular development and maturation have been linked to both temporary and chronic forms of hypothyroidism. Decreased levels of malondialdehyde, increased hydrogen peroxide, increased protein carbonyls, and increased mitochondrial lipid peroxidation are all indicators of oxidative stress in hypothyroid rats. In such situations, testicular mitochondrial protein carbonylation is frequently employed as an indication of oxidative stress [70]. Additionally, the ratio of reduced to oxidized glutathione in the testes is lowered in hypothyroid rats due to decreased GSH levels and increased oxidized glutathione.
It has been demonstrated that in immature rat testes, persistent hypothyroidism disrupts proper redox homeostasis [70]. There have been reports of decreased glutathione-S-transferase activity, elevated GPx, and decreased SOD and CAT activities. Nonetheless, some research indicates that chronic hypothyroidism may decrease glutathione reductase and GPx levels while increasing SOD and CAT activity [50]. Reduced levels of seleno-dependent and seleno-independent GPx are another effect of chronic hypothyroidism [70], suggesting that SOD and CAT have a greater role in controlling oxidative stress in these circumstances.
Because testosterone's metabolism necessitates protection from peroxidative damage, the decrease in GPx activity has a detrimental effect on testosterone production. Reduced semen quality results from lower serum testosterone levels, which also impair spermatogenesis, increase germ cell death, and decrease testicular germ cell populations [69, 70]. In terms of morphology, hypothyroidism causes the diameter of seminiferous tubules to decrease. These alterations in testicular function brought on by hypothyroidism may eventually show up as decreased adult fertility, with signs of low-quality semen, decreased germ cell viability, and decreased sperm counts [69, 70].
2.4 Crosstalk of thyroid hormones with growth factors and other hormones in regulation of male reproductive functions:
Prolactin at physiological quantities has been demonstrated to directly activate Sertoli cells, increasing the synthesis of androgen-binding protein (ABP), a crucial component of Sertoli cell proliferation and metabolism. Koivisto et al. found that a single intravenous dosage of thyroxine dramatically increased prolactin levels in a model of metoclopramide-induced short-term hyperprolactinemia in male Beagle dogs. Semen parameters, luteinizing hormone (LH), and testosterone levels, however, did not show any appreciable alterations. Significantly greater prolactin levels were seen in hypothyroidism patients than in control subjects (P<0.001) [71].
Evidence suggests that thyroid malfunction may be linked to illnesses like acromegaly and increased growth hormone levels. 42.1% of the 57 acromegaly patients had erectile dysfunction, particularly those with a longer history of the condition [72].
IGF-1 and thyroid hormone (T3) seem to work in concert to promote the growth and differentiation of type A undifferentiated spermatogonia by boosting IGF-1 bioactivity via blocking IGF-binding proteins [72].
Obesity and weight gain are common side effects of hypothyroid disorders, and they are linked to increased adipose tissue production of leptin. Leptin has both central and peripheral effects on reproductive function. Leptin's regulating role in male reproductive health is demonstrated by the discovery of leptin receptors in seminal vesicles, the prostate, and Leydig cells. Although leptin also has indirect effects that may impact GnRH signaling, it has been demonstrated to directly reduce testicular testosterone synthesis. Reduced testicular weight, volume, seminiferous tubule diameter, and spermatocyte count can result from hyperleptinemia, which can also stimulate cytokine signaling-3 and decrease the expression of phosphorylated STAT-3. Leptin and thyroid hormones also interact; research has shown that body mass index, leptin, and serum thyroid-stimulating hormone (TSH) are positively correlated. Elevated leptin levels may be a compensatory mechanism for weight gain brought on by hypothyroidism in hypothyroid people [73].
Thyrotoxicosis, a clinical condition caused by excessive exposure to thyroid hormones, is most commonly caused by hyperthyroidism, which is most often caused by Graves' disease (GD), an autoimmune ailment, during the reproductive years. About 0.2% of women experience GD during pregnancy, and 0.4% to 1.0% do so before [74]. It is crucial to distinguish GD from hyperemesis gravidarum, which affects 0.3–1% of pregnancies and is a more prevalent illness. Subacute thyroiditis, toxic adenoma, and toxic multinodular goiter are less common causes, but additional etiologies are extremely uncommon. For the right treatment to be administered, the underlying cause must be accurately identified. Compared to GD, gestational transitory thyrotoxicosis is more prevalent. TSH receptor gene abnormalities that cause increased sensitivity to hCG are an uncommon cause of hyperthyroidism during pregnancy. Maternal TSH levels frequently decrease throughout the first trimester, particularly around the hCG peak between weeks 7 and 11 of pregnancy, since hCG can trigger TSH receptors. By week 11, up to 5% of pregnant women may have TSH levels as low as 0.1 mU/L [75].
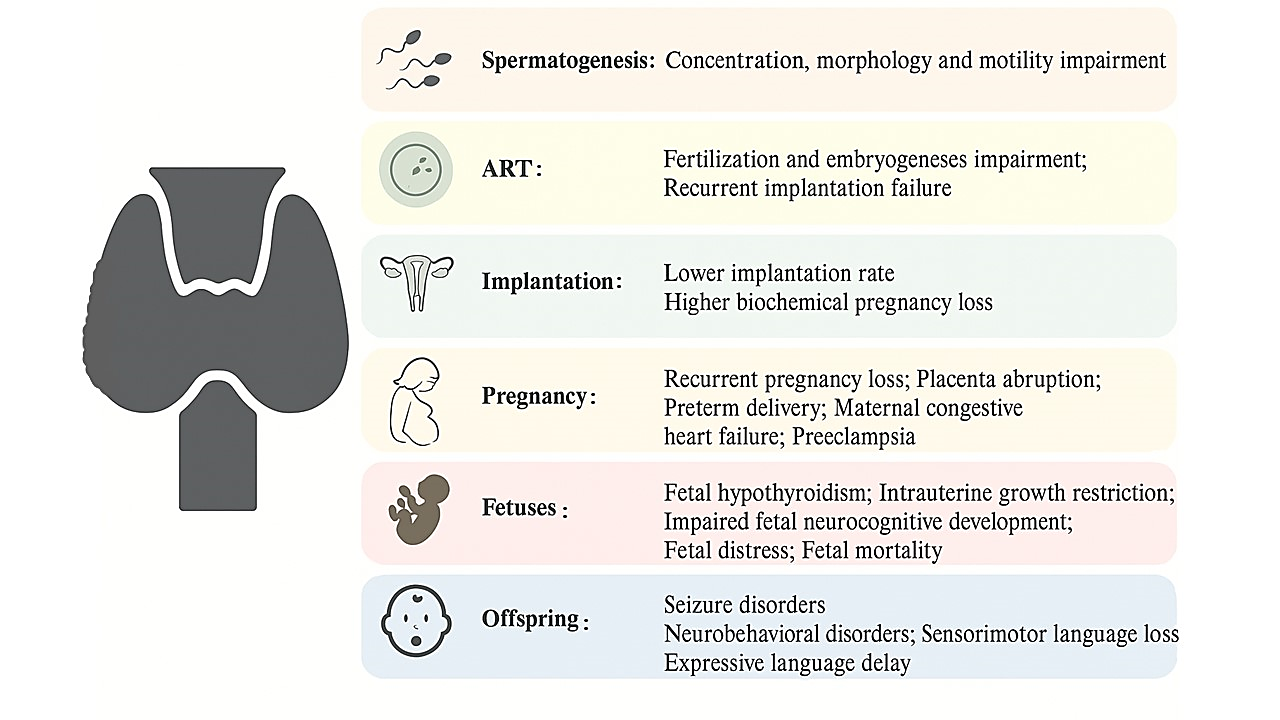
Figure 5: Representation of how thyroid dysfunction impacts reproductive physiology in both males and females.
In the general population, the overall prevalence of overt and subclinical hypothyroidism is between 0.3-0.4% and 4.3-8.5%, respectively. The primary causes of these events are drug usage, autoimmune illnesses, radiation exposure, and thyroidectomy. Prevalence estimates for overt and subclinical hypothyroidism during pregnancy are 0.3-0.5% and 2-3%, respectively, with the latter occasionally reaching 5% [75]. The primary causes of hypothyroidism during pregnancy in iodine-sufficient regions are endemic iodine deficiency and autoimmune thyroiditis [81]. Although there is ample evidence linking overt hypothyroidism to infertility and unfavorable pregnancy outcomes, the effects of subclinical hypothyroidism are less clear. There is significant discussion and critical evaluation about the proper reference ranges for thyroid-stimulating hormone (TSH) levels in connection to assisted reproduction, pregnancy, and fertility.
Karmon et al. observed that preconception TSH levels between 2.5 and 4.9 mIU/L did not correspond with unfavorable clinical or obstetric outcomes, such as a lower live birth or a higher rate of miscarriage, when assessing the results of intrauterine insemination (IUI) [86]. Additionally, groups with TSH values between 2.5 and 5.0 mIU/L and those with TSH values below 2.5 mIU/L did not differ statistically significantly in live birth, pregnancy, or miscarriage rates, according to Unuane et al. [87]. It's critical to acknowledge the significant variation across current research. The main cause of this is the changing definitions of "normal thyroid function" and "subclinical hypothyroidism," which have an impact on study interpretations and treatment choices. Conclusions are made more difficult by discrepancies in study design, outcome definitions, and reporting guidelines. A patient's age, body mass index, history of IVF, and cause of infertility all have a big impact on the results. Results may also be impacted by the particular IVF procedures and fertilization techniques (such as ICSI for male factor infertility) and the quantity of embryos transferred. When taken as a whole, these factors make it challenging to develop consistent clinical recommendations.
According to a number of studies, a subclinical form of hypothyroidism may be associated with an increased risk of miscarriage, preterm birth, preeclampsia, increased fetal mortality, and developmental delays in children's neuropsychological functions and vision, depending on the threshold for TSH used to define the condition [82]. Furthermore, studies have shown that women with TSH levels less than 2.5 mU/L have much reduced miscarriage rates than those with levels greater than 2.5. In a large research of 17,298 pregnant women, Casey et al. established a relationship between subclinical hypothyroidism (TSH > 3) and an increased risk of premature birth (before 34 weeks) [89]. Cleary-Goldman et al., however, did not support these findings, finding no connection between high TSH and preterm delivery (before 37 weeks) [90].
Levothyroxine (L-T4) was used to treat biochemical hypothyroidism in 44 men in a research by Krassas et al. The SHIM questionnaire was used both before and one year after the patients reached a euthyroid condition. Originally seen in 84.1% (37 out of 44), a SHIM score of 21 or below indicated a degree of erectile dysfunction (ED). 29.5%, were still in this range. Following treatment, there was a significant improvement in SHIM scores (mean score increased from 17.0 to 24.0; P < 0.0001). A year later, treated patients' SHIM ratings did not change statistically from those of healthy controls [100]. L-T4 was used in a similar manner by Carani et al. to restore euthyroidism in 14 hypothyroid men. Significant decreases in intravaginal ejaculation latency time (IELT) were observed in half of the participants, along with a resolution of delayed ejaculation (DE). Interestingly, IELT dropped after therapy whether or not DE existed at baseline. The prevalence of ED dropped from 64.3% (9 out of 14) to 21.4% (3 out of 14). Intercourse satisfaction was the only individual dimension with a statistically significant change, despite the fact that IIEF ratings improved significantly. Following treatment, patients with hypoactive sexual desire disorder (HSDD) also reported significant symptom alleviation [66]. Sexual function in women was similarly positively impacted by thyroid hormone supplementation. According to Oppo et al., hypothyroid women who received treatment to attain euthyroidism saw complete normalization in the FSFI domains of pain, desire, and satisfaction, but only slight changes in the arousal and orgasm domains [101].
Additionally, 27 hyperthyroid individuals on antithyroid medications such methimazole were examined by Krassas et al. Following treatment, the number of patients with SHIM scores indicating ED (≤21) decreased to 25.9% (7 of 27), down from 70.4% (19 of 27) prior to treatment. After reaching euthyroidism, SHIM scores significantly increased (mean score increased from 14.5 to 23.0; P<0.0001) [100]. After thyroid function returned to normal, 34 hyperthyroid males participated in another trial that shown significant improvements in IIEF scores, especially in the areas of erectile function and intercourse satisfaction. Premature ejaculation (PE) prevalence decreased from 50% to 15%, which is the predicted rate in the general population. The majority of subjects in the study also showed improvement or resolution of both DE and HSDD [66]. Thyroid hormone normalization dramatically decreased the prevalence of severe ED from 28.6% to 0%, according to additional research by Corona et al. [102]. Cihan et al. treated 24 hyperthyroid men with radioiodine (n = 7), thyroidectomy (n = 7), or antithyroid medication alone (n = 10). After treatment, the percentage of patients having definite PE, as determined by IELT, decreased from 66% (16 of 24) before to 25% (6 of 24). After therapy, the IELT also increased dramatically, going from 75.8 ± 99.3 seconds to 123 ± 96.4 seconds. All IIEF domains showed improvements as well, with the exception of intercourse satisfaction [103]. Similar to hypothyroidism, hyperthyroid women's sexual function improved in a number of ways after their thyroid levels returned to normal. Returning to a euthyroid condition considerably enhanced FSFI dimensions like desire, arousal, lubrication, satisfaction, and pain in a study involving 22 women; however, the orgasm domain showed no discernible changes. Interestingly, pain scores were still greater than those of healthy euthyroid controls even after improvements [101].
The intricate relationship between thyroid function and human reproductive health underscores the essential role of thyroid hormones in maintaining sexual function and fertility across both sexes. Dysregulation of thyroid hormones-manifesting as either hypothyroidism or hyperthyroidism-has been consistently associated with a range of reproductive abnormalities. In men, thyroid dysfunction alters spermatogenesis, affects seminal parameters, and contributes to erectile and ejaculatory dysfunction. In women, thyroid disorders influence menstrual regularity, ovarian reserve, and contribute to infertility, miscarriage, and adverse pregnancy outcomes. Both genomic and non-genomic pathways mediate these effects, involving thyroid hormone receptors, transport proteins, and a network of endocrine and paracrine signals that interact with the hypothalamic-pituitary-gonadal axis.
Despite the accumulating evidence, several gaps persist. For instance, while the physiological influence of thyroid hormones on reproductive tissues is well described, the precise molecular mechanisms-especially those underpinning sex-specific effects-remain poorly elucidated. Furthermore, the threshold levels at which thyroid hormone deviations begin to significantly impair fertility parameters are not yet well standardized, especially in subclinical cases. Additionally, psychological and metabolic confounders, often present in thyroid disorders, are frequently overlooked when assessing sexual dysfunction.
Future research must focus on large-scale, longitudinal studies to clarify the temporal relationship between thyroid dysfunction and reproductive outcomes. There is also a need for integrative models that can delineate the interplay between thyroid hormones, other metabolic regulators (e.g., leptin, insulin), and inflammatory mediators in the context of reproductive dysfunction. The development of more precise biomarkers to predict reproductive risk in thyroid disease patients could enhance early intervention strategies.
Clinically, a multidisciplinary approach integrating endocrinology, andrology, gynecology, and mental health services is essential for optimal management. Early screening for thyroid dysfunction in individuals presenting with idiopathic infertility or unexplained sexual dysfunction may uncover reversible causes and improve therapeutic outcomes. Finally, future guidelines should aim to stratify risk based on thyroid hormone levels and reproductive profiles, thereby moving toward more personalized medicine in reproductive endocrinology.
REFERENCES




Mahesh Kumar Chawra*, Devesh Yaduvanshi, Fiza A. Shaikh, Sanjita Oberai, Thyroid Hormones and It's Effects on Sexual Health of Males and Females, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 6, 4093-4118. https://doi.org/10.5281/zenodo.15732835
 10.5281/zenodo.15732835
10.5281/zenodo.15732835
