1Dean, Faculty of Medical Science and Research, Sai Nath University, Ranchi, Jharkhand 835219, India.
2Vice Principal, Faculty of Medical Science and Research, Sai Nath University, Ranchi, Jharkhand 835219, India.
3Associate Professor, Department of Pharmacy, Sai Nath University, Ranchi, Jharkhand 835219, India.
4Student, B. Pharm, Department of Pharmacy, Sai Nath University, Ranchi, Jharkhand 835219, India.
5Assistant Professor of Pharmacology, Department of Pharmacy, Faculty of Medical Science and Research, Sai Nath University, Ranchi, Jharkhand 835219, India
Anxiety disorders, including generalized anxiety disorder (GAD), panic disorder, post-traumatic stress disorder (PTSD), and obsessive-compulsive disorder (OCD), present a significant public health and economic burden globally. Traditional pharmacotherapies, primarily targeting the ?-aminobutyric acid (GABA) and serotonergic systems, are associated with substantial limitations, including treatment resistance and side effects. Emerging research suggests that dysregulation in glutamatergic neurotransmission may play a key role in anxiety pathophysiology, offering novel therapeutic targets. Neuroimaging studies have identified structural and functional alterations in glutamate-rich regions, such as the hippocampus and amygdala, in individuals with anxiety disorders. Moreover, pharmacological modulation of glutamate receptors, including NMDA, AMPA, and metabotropic glutamate receptors, has demonstrated efficacy in reducing anxiety symptoms. Agents such as D-cycloserine (NMDA partial agonist) and riluzole (glutamate release inhibitor) have shown promise in alleviating symptoms of PTSD and OCD. These findings support the potential of glutamate-based therapies as an innovative approach to treating anxiety disorders. Further research is warranted to elucidate the precise mechanisms and optimize the clinical application of glutamate-targeting treatments.
Anxiety disorders, encompassing generalized anxiety disorder (GAD), post-traumatic stress disorder (PTSD), panic disorder, and obsessive-compulsive disorder (OCD), are among the most common and debilitating psychiatric conditions globally [1, 2]. These disorders not only significantly impair individual well-being and daily functioning but also contribute to a substantial economic burden through increased healthcare costs and diminished productivity due to absenteeism and disability. Current pharmacological interventions primarily focus on modulating the ?-aminobutyric acid (GABA) and serotonergic systems, utilizing agents such as benzodiazepines and selective serotonin reuptake inhibitors (SSRIs). However, these treatments are frequently associated with limitations, including partial efficacy, treatment resistance, and adverse side effects such as sedation, cognitive impairment, and the risk of dependency, which can lead to poor adherence and relapse. Emerging evidence has identified the glutamatergic system as a crucial player in the neurobiology of anxiety disorders. Dysregulation in glutamate signaling, particularly within the NMDA (N-methyl-D-aspartate) and AMPA (?-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) receptor pathways, is implicated in the pathogenesis of anxiety-related behaviors. Pharmacological modulation of these receptors presents a novel therapeutic strategy [3-7]. Agents such as D-cycloserine, a partial NMDA receptor agonist, and riluzole, a glutamate release inhibitor, have demonstrated efficacy in attenuating symptoms in anxiety disorders, including PTSD and OCD. These findings highlight the therapeutic potential of targeting glutamatergic transmission, offering a new frontier in the treatment of anxiety disorders that may overcome the limitations of traditional therapies. Continued research is essential to further elucidate the precise mechanisms by which glutamatergic modulation can be optimized to improve clinical outcomes for patients suffering from anxiety-related conditions [8-10].
Mechanistic Role of Glutamate in Anxiety: Receptor Dynamics and Neural Circuitry
Glutamate, the brain's principal excitatory neurotransmitter, plays a vital role in anxiety regulation by influencing key neuronal circuits involved in emotional responses, stress adaptation, and fear processing. The underlying mechanism by which glutamate impacts anxiety is closely linked to its action on both ionotropic and metabotropic glutamate receptors. These receptors include NMDA (N-methyl-D-aspartate), AMPA (?-amino-3-hydroxy-5-methyl-isoxazolepropionic acid), kainate receptors, and various metabotropic glutamate receptors (mGluRs), which are widely distributed across critical brain regions like the amygdala, hippocampus, prefrontal cortex, and anterior cingulate cortex—areas essential for emotional regulation and implicated in anxiety disorders [11-17]. During periods of stress or elevated anxiety, glutamate release tends to increase, resulting in overactivation of these receptors [18, 19]. NMDA receptors, in particular, are crucial for synaptic plasticity, learning, and memory consolidation, including the encoding of fear-related memories [20, 21]. However, chronic overactivation of NMDA receptors can lead to excitotoxicity, causing neuronal damage, which may contribute to the persistence of anxiety in conditions such as post-traumatic stress disorder (PTSD) and generalized anxiety disorder (GAD) [22-24]. Similarly, AMPA and kainate receptors are integral to fast excitatory synaptic transmission and fear responses [25, 26]. Dysregulation of AMPA receptor activity, in particular, has been linked to excessive neural circuit excitation, which manifests as heightened fear responses, hyperarousal, and anxiety-related behaviors [27, 28]. Metabotropic glutamate receptors, notably mGluR2/3 and mGluR5, play an important role in modulating glutamate release and regulating neuronal excitability [29, 30]. Agonists of mGluR2/3 have shown anxiolytic effects by reducing excessive glutamate release, thereby mitigating hyperactivity in anxiety-related brain neurones [31, 32]. This reduction in glutamate signalling helps prevent overstimulation of neural pathways that contribute to anxiety. On the contrary, antagonists of mGluR5 have also demonstrated therapeutic potential in reducing anxiety-like behaviors, likely due to their ability to inhibit glutamate's excitatory effects [33-35]. Chronic dysregulation of glutamate transmission not only alters synaptic activity but also leads to structural changes in the brain. For example, hippocampal atrophy and amygdala hypertrophy, which are frequently observed in individuals with anxiety disorders, exacerbate the cycle of anxiety symptoms. These structural changes reinforce the dysfunction in glutamatergic signaling, contributing to emotional dysregulation. Targeting glutamate signaling pathways offers a promising approach for developing novel treatments aimed at restoring balance in glutamatergic transmission and alleviating the symptoms of anxiety disorders [36-40].
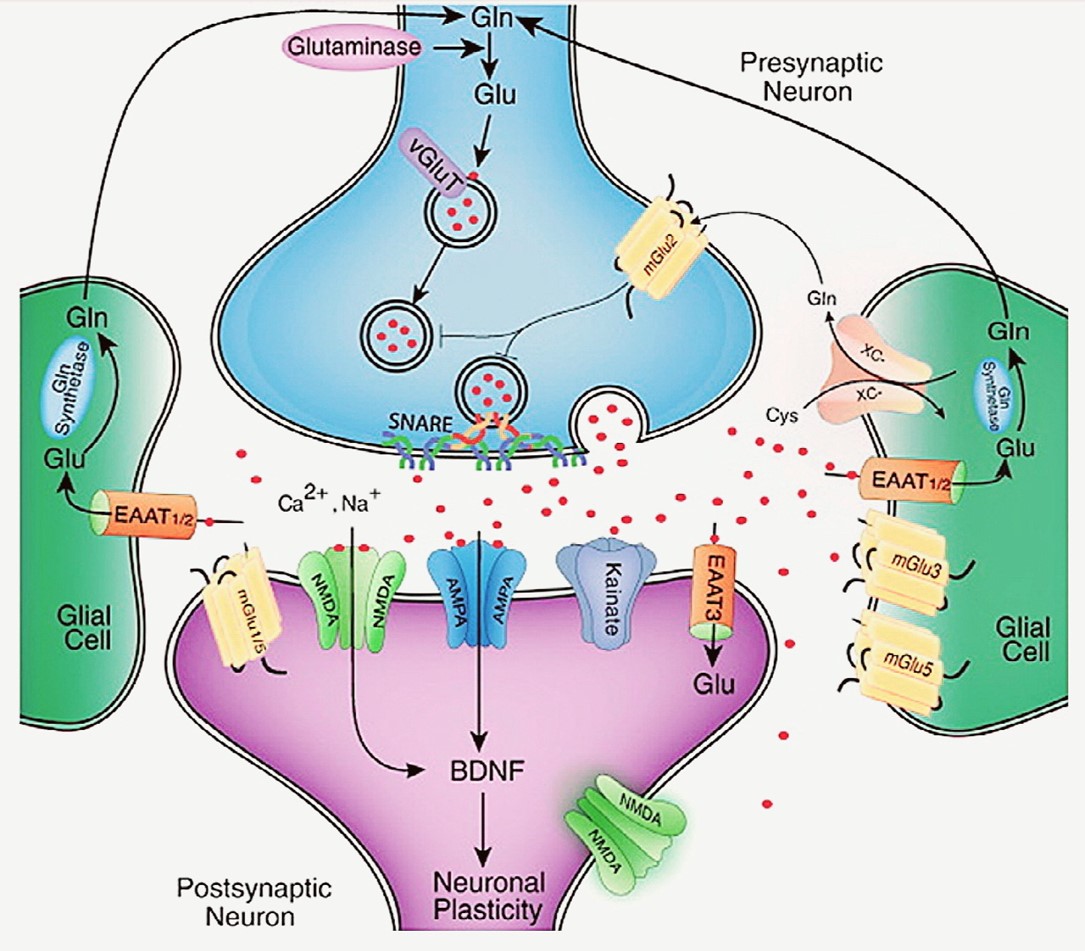
Fig. 1. This is a schematic representation of a glutamatergic synapse which illustrates that the synthesis, release, and regulation of glutamate (Glu), along with its associated receptors and deactivation pathways. Glutamine (Gln), converted to Glu by the enzyme glutaminase, serves as the primary source of synaptic glutamate, though Glu can also be produced from intermediates of the tricarboxylic acid (TCA) cycle. Once synthesized, Glu is transported into presynaptic vesicles by vesicular glutamate transporters (VGluTs) and released into the synaptic cleft via a voltage-dependent mechanism involving SNARE protein complexes. Following its release, glutamate is removed from the extracellular space predominantly by excitatory amino acid transporters (EAATs) located mainly on astroglial cells. In astrocytes, Glu is converted back to Gln by the enzyme glutamine synthetase, and Gln is then exported and taken up by neurons for reuse in synaptic transmission. Another mechanism for Glu recycling involves the system x-C, a cystine/glutamate antiporter on glial cells, which also facilitates Glu reuptake.Glutamate receptors are expressed on both presynaptic and postsynaptic neurons, as well as on glial cells. These receptors are classified into two major categories: ionotropic receptors, including AMPA (?-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid), NMDA (N-methyl-D-aspartate), and kainate receptors, and metabotropic glutamate receptors (mGluRs). The effects of glutamate signaling are determined by the specific receptor subtype and its location within synaptic, perisynaptic, or extrasynaptic sites. Brain-derived neurotrophic factor (BDNF) also plays a role in modulating glutamatergic synaptic function and plasticity.
The Role of Glucocorticoids and Glutamatergic Excitotoxicity in Stress-Induced Neuroanatomical and Behavioural ChangesThe release of adrenal glucocorticoids is a typical physiological response to stress [41, 42]. However, chronic exposure to stress results in prolonged glucocorticoid release, which has been linked to neurotoxic events, including increased glutamate release, particularly in the hippocampus [43, 44]. For example, animal models exposed to repeated immobilization or forced swimming stress show elevated glutamate release and uptake in the hippocampus and prefrontal cortex [45, 46]. Moreover, chronic stress has been associated with altered gene expression, such as increased AMPA receptor mRNA levels in the hippocampus, further emphasizing glutamate's role in stress-induced neurotoxicity [47-50]. Chronic stress-induced glucocorticoid release can lead to hippocampal atrophy, including decreased dendritic branching, neuronal death, and reduced neurogenesis in hippocampal pyramidal cells [51, 52]. Conversely, stress exerts hypertrophic effects in the amygdala, promoting increased dendritic arborization in pyramidal and stellate neurons of the basolateral amygdala and bed nucleus of the stria terminalis [53, 54]. These paradoxical anatomical changes reflect the contrasting roles of the hippocampus and amygdala in stress circuitry, with the hippocampus exerting inhibitory effects on the hypothalamic-pituitary-adrenal (HPA) axis, while the amygdala promotes excitatory regulation [55, 56]. Behavioural studies highlight the divergent effects of stress on hippocampal- and amygdala-dependent learning. Chronic stress impairs hippocampal-dependent spatial learning, as evidenced by deficits in tasks like the radial arm maze and Morris water maze, both of which rely on glutamatergic signalling [57, 58]. In contrast, glutamate release in the amygdala enhances fear-related learning, such as contextual fear conditioning. Pharmacological studies underscore the importance of NMDA receptors in both the hippocampus and amygdala for the acquisition and expression of fear-related memories, suggesting that glutamatergic pathways play a key role in emotional processing under stress. This mechanism is further implicated in disorders like PTSD, where glutamatergic dysregulation may contribute to symptoms such as dissociation and altered perception, similar to the effects of NMDA receptor antagonists like ketamine [59-63]. Various pharmacological agents targeting glutamate receptors and associated pathways have demonstrated significant effects on fear-potentiated startle and other anxiety-related behaviors. NMDA receptor antagonists, such as AP5, have been shown to reduce fear-potentiated startle, while NMDA partial agonists, like D-cycloserine (DCS), exhibit mixed effects, either enhancing or reducing responses depending on the specific behavioral paradigm. For example, DCS reduces fear-potentiated startle but enhances cue-conditioned freezing during extinction. It also affects behaviors in the elevated plus maze (EPM), with both increases and decreases in time spent in open arms, depending on the study [64, 65].
The Role of Glutamate Receptor Agonists and Antagonists in Modulating Stress-Induced Behaviours
Several agents targeting AMPA/kainate receptors exhibit similar variability. Kainic acid, an agonist, decreases fear-potentiated startle, while antagonists like NBQX increase it. AMPA/kainate receptor antagonist LY326325 increases punished drinking and time spent in open arms during the EPM [66, 67]. Topiramate, an AMPA/kainate receptor agonist, reduces stress-induced startle, highlighting its anxiolytic potential [68, 69]. In the metabotropic glutamate receptor (mGluR) category, mGluR5 antagonists, such as MPEP, decrease fear-potentiated startle but increase punished responding, suggesting complex modulation of anxiety behaviors [70, 71]. mGluR2/3 agonists like LY354740 reduce fear-potentiated startle and panic-like responses, such as lactate-induced panic, while also increasing time spent in open arms during the EPM [72, 73]. Additional agents, including sodium channel blockers (e.g., lamotrigine) and glutamate release inhibitors (e.g., riluzole), enhance conditioned emotional responses, indicating their potential in modulating anxiety-related behaviors through glutamatergic mechanisms. These findings collectively demonstrate the diverse and context-dependent effects of glutamate receptor modulation on stress and anxiety responses [74-78].
In addition to this, different pharmacological agents targeting glutamate signalling have shown promise in reducing symptoms associated with various anxiety disorders and phobias [79, 80]. The mGluR2/3 agonist LY354740 has been found to reduce fear-potentiated startle and panic responses during CO2 challenge, while LY544344, another mGluR2/3 agonist, has been shown to alleviate panic responses to CCK-4 challenge [81, 82]. Phenytoin, which inhibits glutamate transmission, has demonstrated a decrease in symptoms of PTSD [83, 84]. Similarly, the NMDA partial agonist D-cycloserine (DCS) has been associated with reductions in PTSD symptoms and specific phobias [85, 86]. Topiramate, an AMPA/kainate receptor agonist, has been shown to decrease symptoms of PTSD and social phobia in multiple studies [87, 88]. Additionally, Riluzole, a glutamate release inhibitor, has shown efficacy in reducing symptoms of obsessive-compulsive disorder (OCD) and generalized anxiety disorder (GAD). These findings support the role of glutamatergic modulation in the treatment of stress and anxiety-related disorders [89, 90].
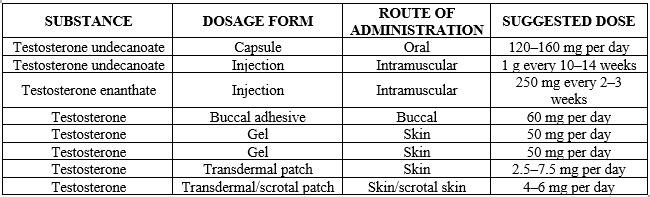
Table-I. Pharmacological Agents and Their Effects on Anxiety-Related Behaviours
The Role of Glutamate in Anxiety Disorders: Genetic, Clinical, and Pharmacological Evidence
Although limited, human genetic, physiological, and behavioral studies provide preliminary evidence supporting the involvement of the glutamate system in fear and anxiety. Research has identified a significant association between the NMDA receptor subtype 2B (GRIN2B) gene and obsessive-compulsive disorder (OCD), with a specific single nucleotide polymorphism (5072T/G) linked to increased OCD risk and symptom severity. Elevated cerebrospinal fluid (CSF) glutamate levels have also been observed in patients with OCD, adding to the evidence of glutamatergic involvement in anxiety disorders, though the exact relationship between brain and CSF glutamate remains unclear [91-95]. Pharmacological studies further underscore the role of glutamate modulation in anxiety. The mGluR2/3 agonist LY354740 has demonstrated anxiolytic effects, reducing fear-potentiated startle in humans, and its prodrug LY544344 has shown promise in reducing anxiety induced by cholecystokinin tetrapeptide (CCK-4). Moreover, clinical trials targeting glutamate receptors have shown efficacy in treating anxiety disorders such as PTSD, phobias, and OCD. Phenytoin, which reduces glutamate transmission, significantly decreased PTSD symptoms, while D-cycloserine (DCS), an NMDA partial agonist, has been shown to enhance the extinction of fear in patients with acrophobia and PTSD [96-100].
Non-NMDA glutamate receptors also play a role in anxiety. Topiramate, an AMPA/kainate receptor modulator, has shown effectiveness in treating chronic PTSD and social phobia, likely due to its dual modulation of both excitatory (glutamate) and inhibitory (GABA) neurotransmission. Similarly, riluzole, a glutamate release inhibitor, has demonstrated efficacy in treating generalized anxiety disorder (GAD) and treatment-resistant OCD, with promising results in early clinical trials [101-103].
These findings highlight the significant role of glutamate receptor modulation in the treatment of anxiety disorders and suggest that further research into glutamatergic mechanisms could lead to novel therapeutic approaches for these conditions [104, 105].
Glutamate's Role in Anxiety Disorders: Insights from Neuroimaging Studies
Magnetic resonance imaging (MRI) has provided significant insights into the relationship between glutamate and anxiety disorders, albeit indirectly. Structural MRI (volumetric) and functional MRI (fMRI) studies have shown that glutamate-rich brain regions, such as the hippocampus, amygdala, and anterior cingulate cortex (ACC), are often structurally or functionally altered in individuals diagnosed with anxiety disorders [106, 107]. For instance, trauma survivors with post-traumatic stress disorder (PTSD) show reduced gray matter volume in the anterior cingulate and hippocampus. Moreover, studies have shown that combat veterans with PTSD, as well as women with PTSD related to childhood abuse, exhibit reduced hippocampal volume [108, 109]. Although not all studies consistently report hippocampal changes in PTSD, a meta-analysis has confirmed smaller hippocampal volumes in individuals with PTSD compared to healthy and traumatized controls [110]. This reduction in hippocampal volume is consistent with findings from animal studies showing stress-induced atrophy in the hippocampus [111]. Interestingly, in contrast to the hippocampus, the amygdala demonstrates hypertrophy in children and adolescents diagnosed with anxiety disorders, such as generalized anxiety disorder (GAD) and obsessive-compulsive disorder (OCD) [112, 113]. These structural alterations align with functional brain imaging studies, such as positron emission tomography (PET) and fMRI, which have shown altered activity in these regions. For example, patients with panic disorder exhibit abnormal metabolism in the hippocampus and ACC, while veterans with PTSD show altered blood flow in the amygdala [114-116]. Furthermore, fMRI studies have linked hyperactive amygdala responses to negatively biased social cues in individuals with social anxiety disorder, highlighting the role of glutamatergic pathways in emotional processing [117, 118]. Proton magnetic resonance spectroscopy (1H-MRS) has allowed researchers to directly quantify neurochemicals, including glutamate, in vivo. Although much of the focus has been on N-acetylaspartate, recent studies have revealed region-specific changes in glutamate levels in individuals with anxiety disorders [119, 120]. For instance, increased glutamate levels have been detected in the frontal cortex of healthy individuals with higher state-trait anxiety. In patients with social anxiety disorder, elevated glutamate concentrations in the ACC correlate with symptom severity [121, 122]. Similarly, children with OCD exhibit reduced glutamate concentrations in the ACC and elevated glutamatergic activity in the caudate nucleus, with symptom improvement following treatment [123, 124]. These findings suggest a complex tonic-phasic dysregulation of the glutamate system in anxiety, supporting the role of glutamatergic mechanisms in the pathophysiology of anxiety disorders [125].
Economic and Public Health Burden of Anxiety and Related Disorders
Anxiety, stress, and trauma-related disorders present a significant public health challenges in all over the world. This cost is driven by both direct healthcare expenditures, including psychiatric and non-psychiatric services, emergency care, hospitalization, and medication, as well as indirect costs such as reduced workforce productivity and occupational absenteeism. These disorders impose a considerable burden not only on the healthcare system but also on the economy as a whole, underscoring the importance of addressing the needs of affected individuals [126-128].
Prevalence and Associated Comorbidities
Anxiety is a common psychological response that, in normal circumstances, serves an adaptive function by helping individuals manage stress. However, for more than 15 million adults in the U.S. each year, this response becomes maladaptive, resulting in the development of anxiety disorders. These disorders, which include conditions such as generalized anxiety disorder (GAD), social and specific phobias, panic disorder, post-traumatic stress disorder (PTSD), and obsessive-compulsive disorder (OCD), are the most prevalent mental health issues in the U.S. Individuals with anxiety disorders frequently experience comorbid conditions such as irritable bowel syndrome (IBS) and hypertension, and they are at increased risk for mood disorders, including depression. These comorbidities compound the impact of anxiety disorders on overall health and well-being [129-132].
Limitations of Current Therapies and the Need for Alternative Approaches
The significant limitations of current treatment options for anxiety disorders, including issues of noncompliance and treatment resistance, highlight the urgent need for novel therapeutic strategies. Emerging research in neuroscience has identified the glutamate system, the brain’s principal excitatory neurotransmitter, as a potential target for new anxiety treatments. Many anxiety and stress-related disorders are thought to arise from hyperactive or overly responsive neural circuits, suggesting that interventions aimed at modulating glutamatergic function could offer a more effective approach to managing these conditions. Investigations into glutamate-modulating drugs hold promise for improving outcomes in individuals with anxiety disorders and advancing our understanding of the underlying mechanisms driving these severe and debilitating mental health conditions [133-135].
CONCLUSION:
In conclusion, anxiety disorders represent a major challenge both in terms of individual health and global public health due to their high prevalence, comorbidities, and significant economic impact. Traditional treatments targeting the GABAergic and serotonergic systems have shown efficacy but are often limited by treatment resistance and adverse effects, underscoring the need for novel therapeutic strategies. Emerging research into the glutamatergic system provides promising insights, with glutamate modulation offering a potential pathway for more effective treatments. Agents like D-cycloserine, riluzole, and others targeting NMDA, AMPA, and metabotropic glutamate receptors have shown encouraging results in alleviating anxiety symptoms, particularly in conditions such as PTSD and OCD. Furthermore, neuroimaging studies reveal structural and functional abnormalities in glutamate-rich brain regions, such as the hippocampus and amygdala, in individuals with anxiety disorders, further supporting the role of glutamatergic dysregulation in the pathophysiology of these conditions. The accumulating genetic, clinical, and pharmacological evidence points to glutamate-based therapies as a potential frontier in the treatment of anxiety disorders. However, more research is required to elucidate the precise mechanisms of glutamatergic modulation and optimize the clinical application of these treatments. By advancing our understanding of glutamate's role in anxiety and refining glutamate-targeting pharmacotherapies, there is hope for more effective and tailored interventions that address the limitations of current therapeutic approaches, ultimately improving the quality of life for those affected by anxiety disorders
REFERENCE
135. Mewton L, Smith J, Rossouw P, Andrews G. Current perspectives on Internet-delivered cognitive behavioral therapy for adults with anxiety and related disorders. Psychology research and behavior management. 2014 Jan 30:37-46



















K. Rajeswar Dutt , Ankita Singh , Mahesh Kumar Yadav , Kristy Kumari , Vishal Kumar , Vicky Kumar , Rishu Raj , Chandan Kumar , Megha Chattaraj , Priyanka Daniel , Manish Kumar Singh , Dhananjay Sahu , Md. Danish Gayas Ansari , Adarsh Kumar Singh , Faiz Alam , Sandeep Kumar , Arnab Roy, Targeting Glutamate Signalling In Anxiety Disorders: Novel Insights And Therapeutic Strategies, Int. J. of Pharm. Sci., 2024, Vol 2, Issue 9, 1453-1469. https://doi.org/10.5281/zenodo.13854617
 10.5281/zenodo.13854617
10.5281/zenodo.13854617
