College of Pharmaceutical Sciences, GMC, Kannur, 670503.
Millions of people worldwide suffer from chronic wounds, such as pressure sores, diabetic foot ulcers, and venous leg ulcers. Microbial biofilms, which are bacterial communities encased in extracellular matrices they produce on their own and that withstand host immune responses and antimicrobial treatments, are the main cause of these wounds' delayed healing and high recurrence rates. With a focus on cutting-edge natural treatment approaches, this review investigated biofilm formation mechanisms, resistance patterns, and persistence in chronic wounds. Bromelain, a proteolytic enzyme from pineapple (Ananas comosus), was highlighted in a thorough literature review that concentrated on recent developments in biofilm disruption. Pseudomonas aeruginosa and Staphylococcus aureus, two important biofilm-forming pathogens, have a high level of resistance to antibiotics and prolong chronic inflammation. Natural products hold significant promise in enhancing antimicrobial efficacy and disrupting biofilm structures. Bromelain enhances antibiotic penetration, promotes enzymatic debridement, and demonstrates broad-spectrum antibacterial and antibiofilm qualities. When used against mature biofilms, especially MRSA strains, the bromelain-N-acetylcysteine combination (BromAc) showed remarkable efficacy. While many natural substances exhibit antibiofilm properties, such as tea tree oil (which works against Candida albicans), tannic acid (which targets E. coli CsgD regulators), usnic acid (which inhibits P. aeruginosa adhesion), and ?-amylase (which disrupts extracellular matrices), bromelain stands out due to its special combination of matrix degradation and increased antibiotic permeability, according to comparative analysis. Synergistic combinations, such as flogomicina (NAC, bromelain, and plant extracts), increase the effectiveness of antibiotics and reduce bacterial growth by more than 80%. Although more comparative research is required to completely clarify its clinical potential in the management of chronic wounds, bromelain's triple therapeutic action proteolysis, antibiofilm activity, and wound healing promotion makes it a strong contender for both standalone and combination therapies.
Chronic wounds are a rising worldwide health problem, with an estimated 6.5 million United States patients alone and an annual cost of treatment of more than $25 billion. Such wounds failure to heal within 4-6 weeks with or without chronic wounds such as diabetic foot ulcers, pressure ulcers, and venous leg ulcers that all contribute to patient morbidity, healthcare cost, and prolonged hospital stay.(1) one of the most significant factors preventing chronic wound healing is the development of Microbial biofilms, which are aggregate colonies of bacteria in a self-produced matrix of extracellular polymeric substances (EPS).(2) these biofilm provide protective niches that cover microorganism from antimicrobial agents and host immune cells, significantly prevent infection control and hindering wound closure.(3) Although biofilms are found in acute wounds, they are common in chronic wounds(4) ), tend to be polymicrobial communities, such as both Gram-positive and Gram-negative bacteria, such as Escherichia coli, Pseudomonas aeruginosa, and Staphylococcus aureus.(5)
Polysaccharides, proteins, lipids, and extracellular DNA (eDNA) EPS matrix contribute to structural stability and increased bacterial adhesion and in addition creates diffusion barrier that prevents penetration by antibiotics and clearance by immune cells.(6) ) The architectural sophistication makes standard treatments such as topical antibiotics and mechanical debridement inadequate for total eradication of biofilm.(7)
Current studies have reviled promising antibiofilm candidates acting against multiple disruption mechanisms: quorum sensing inhibition, matrix-degrading enzymes, and biofilm disassembly agent(8) Natural bioactive compounds such as usnic acid, tannic acid, bromelain, and tea tree oil have shown great potential to biofilm structure modification and augmentation of conventional treatment efficacy(9) (10) Of these drugs, bromelain, a mixture of proteolytic enzymes isolated from the pineapple (Ananas comosus), has gained prominence due to its potent antimicrobial, anti-inflammatory, and enzymatic debridement activities. Beyond inhibiting the structure of biofilms through proteolytic activity, bromelain enhances the absorption of antibiotics and facilitates tissue regeneration, making it a highly potential agent for the treatment of chronic wounds.(11) The pathophysiology of wound-related biofilms, their resistance mechanisms, and new antibiofilm therapies are discussed below, with specific regard to the therapeutic potential of bromelain in the treatment of chronic wounds.
MICROBIAL BIOFILM
Bacterial biofilms are complex, structured microbial communities where microbial cells are encased in a matrix of extracellular polymeric substance.(2) (3) “Biofilm is the unique pattern of growth in the life cycle of microbes that provides specific properties, advantages and higher level of organization to the free living bacterial cells during colonization”(12) Different bacterial species can develop distinct biofilm architectures even under the same environmental conditions, which is both species specific and dependent on environmental condition.(13)
The following steps make up the biofilm growth process: (1) microorganisms binding to a surface or to one another; (2) microcolony creation; (3) biofilm maturation; and (4) biofilm dispersal (13). The organic polymers of microbial origin known as extracellular polymeric substances (EPS) are engaged in the interactions between bacterial cells and their surroundings. “a strong and sticky framework”. EPS is a non-cellular three-dimensional structure made up of proteins, glycoproteins, proteoglycans, and exopolysaccharides.(14)
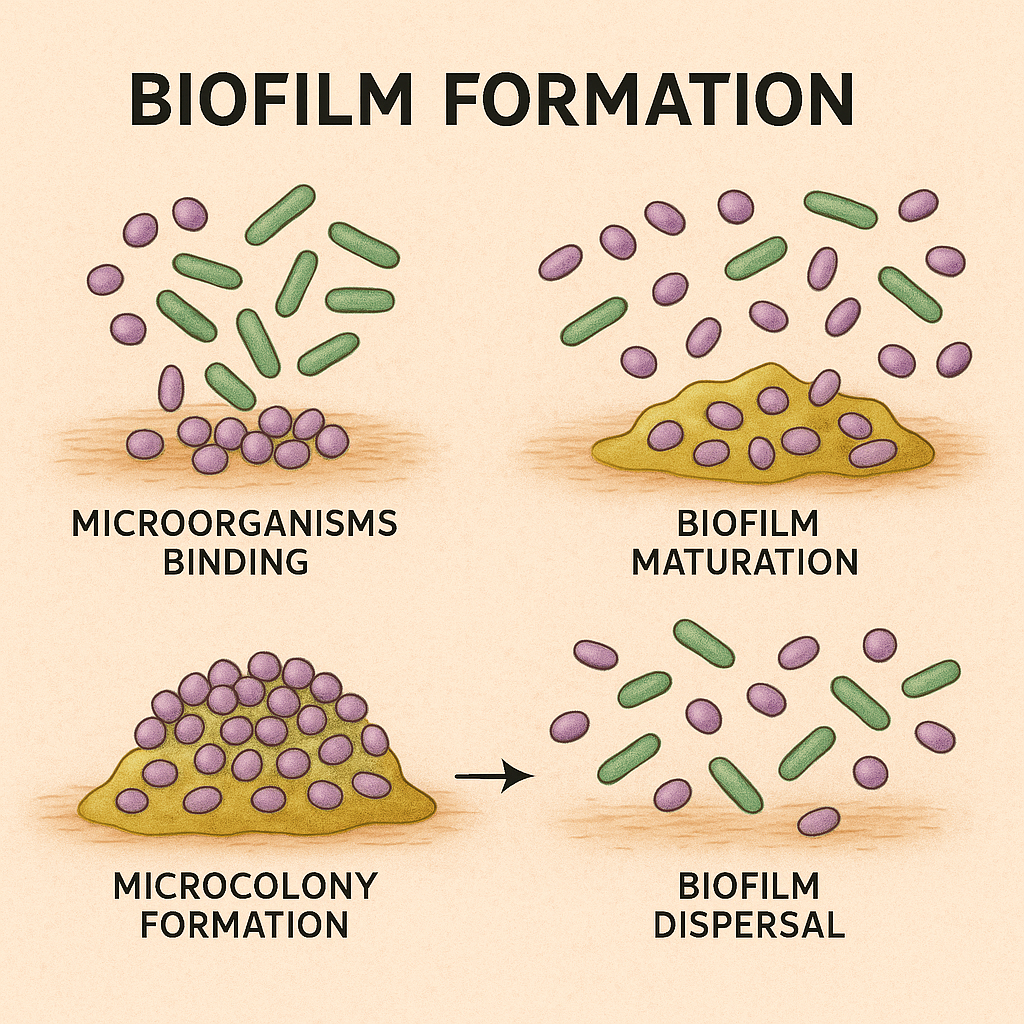
Figure 1: Biofilm growth process- 1. Microorganism binding- attachment of free-floating bacteria to the surface. 2. Microcolony formation- proliferation of attached cells and begin forming clusters. 3. Biofilm maturation- development of a complex 3D structure with an extracellular polymeric matrix. 4. Biofilm dispersal- some cells detach and revert back to a planktonic state, enabling colonization of new cities.
Wound biofilm
Chronic wounds are those with a tendency to last longer than four to six weeks and do not heal through the normal phases of healing in the normal time frame. They tend to be linked to diseases including diabetes, ischemia, infection, or the creation of microbial biofilms.(4) (15) It is challenging to treat them with standard therapy since they have a tendency to stay in a chronic inflammatory process.
To confirm this statement, microscopic analysis of chronic wound samples has identified tightly aggregated colonies of bacteria covered by an extracellular matrix, a feature of biofilms. These morphological features are strong evidence for the presence of biofilms in chronic wounds. Acute wound samples, however, rarely show such structures, which confirms that biofilms are comparatively infrequent in acute wound pathology.(4)
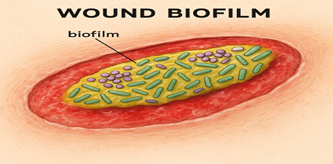
Figure 2: wound biofilm- a mature biofilm with in wound bed.
The resistance and persistence of biofilms also depend on their maturity; Biofilm resistance to disinfectants enhanced by age of biofilm. (16) In an in-vivo model, Pseudomonas aeruginosa biofilm infections were found to cause a significant delay in wound healing by an average of 2 to 4 weeks compared to uninfected controls, indicating the central role of biofilms in the pathophysiology of chronic wounds and the need for effective anti-biofilm therapies.(17) Among the most serious pathogens, Staphylococcus aureus is a main cause of biofilm formation (18) One of its principal surface proteins, designated as Bap (biofilm-associated protein), has 2,276 amino acids and is involved in adhesion and biofilm formation in a wide range of bacterial species. Bap has also been found to enable specific higher intramammary adherence and continuous biofilm formation, as well as the persistence of S. aureus infections.(19) Staphylococcus aureus isolates with a virulent potential to form biofilms are isolated from chronic wounds. (20) Methicillin-resistant S. aureus (MRSA) isolates were highly resistant to clindamycin, erythromycin, and penicillin G, whereas methicillin-sensitive S. aureus (MSSA) isolates commonly showed resistance to penicillin G, cotrimoxazole, and ciprofloxacin. Furthermore, Gram-negative bacteria from chronic wounds have been highly resistant to a number of antibiotics, including amoxicillin-clavulanic acid, ceftazidime, ciprofloxacin, cefoxitin, cephalothin, and cefuroxime. These findings emphasize the clinical challenge posed by drug-resistant biofilm-associated bacteria in the treatment of chronic wounds.
Compared to their non-biofilm-producing cousins, biofilm-producing bacteria are significantly more resistant to antibiotics and disinfectants and therefore harder to treat (21) ) This is confirmed by a study compared mupirocin cream and triple antibiotic ointment for the treatment of Staphylococcus aureus infections. Both were effective against planktonic (free- floating) bacteria but significantly less effective against mature biofilms, despite identical initial bacterial numbers. These findings refer to the reduced susceptibility of biofilm- associated infections and highlight the therapeutic challenge posed by biofilm-producing pathogens. (22) Additionally, polymicrobial biofilms recovered slowly compared to mono-species infection due to increased microbial synergy and increased antimicrobial tolerance. (5) Scanning electron microscopy illustrates that even the most efficient wound care regimen cannot completely eradicate biofilm in a single session, proving biofilm resistance and the need for extensive and sustained treatment. (7)
This issue is not exclusive to only wound infections, Biofilm formation is the single most important cause of device-associated infections, particularly central venous catheter (CVC) associated infections. These catheter-related infections, primarily caused by biofilm-forming bacteria. About 90% of staphylococcal species, including S. epidermidis, S. aureus, and others, produce biofilms. Apart from gram-negative bacteria like E.coli, K. pneumoniae, P. aeruginosa, Candida albicans has a 100% rate of biofilm prevalence, indicating the central role of biofilms in catheter infections.(23) One of the most helpful first-line therapies for the management of biofilm burden in chronic wounds is debridement. The procedure entails the removal of infected or dead tissue and foreign bodies from the wound bed to a level when healthy tissue surrounding the wound becomes visible,(24) is applicable to any lesion, wherever it is from or whatever it is diagnosed as. There are a number of types of debridement Autolytic Debridement, Biological Debridement, Enzymatic Debridement, Surgical Debridement with Sharp Instruments, Mechanical Debridement.
TABLE 1: KEY CHARACTERISTICS OF TRADITIONAL DEBRIDEMENT METHODS IN WOUND CARE
|
Feature |
Debridement |
|
Invasiveness |
Invasive (surgical, mechanical, or enzymatic). |
|
Pain & Discomfort |
Can be painful; may require anesthesia. |
|
Target Specificity |
Non-specific removal of both biofilm and necrotic tissue. |
|
Application Frequency |
Repeated procedures can cause damage to healing tissue. |
|
Use in High-Risk Patients |
May not be suitable for immunocompromised or fragile patients. |
|
Prevention Capability |
Cannot prevent biofilm formation; reactive approach |
|
Tissue Preservation |
Risk of damaging granulation tissue |
|
Ease of Use |
Requires trained personnel or clinical setup |
Although the mechanical method remains a critical strategy, molecular approaches have also evidenced potential in interfering with biofilm formation.
ANTIBIOFILM
The term "antibiofilm" is used to describe any substance or material that blocks or disrupts the formation and growth of microbial biofilms. A wide range of antibiofilm agents has been identified. Numerous compounds, including certain natural extracts, synthetic compounds, enzymes, peptides, chelating agents, polyphenols, and other antibiotics, have been reported to possess anti-biofilm action. Anti-biofilm agents appear to act through different mechanisms against different bacterial species to block biofilm formation.
MECHANISMS OF BIOFILM DISRUPTION
Biofilms in chronic wounds have a high degree of resistance to antimicrobial agents, owing to their well-developed structure and adaptive processes. Interference with these processes using natural antibiofilm agents has been proposed as a potential approach. A number of different modes of action have been described.
QOURAM SENSING INHIBITORS (QSI)
Quorum sensing is a communicative process employed by bacteria to quantify their cell density through chemical signals. When the concentration of the signal surpasses a certain threshold, it triggers group activities such as biofilm formation, making infections difficult to treat.(25)
N-acyl homoserine lactone synthetic analogs have the potential to interfere with quorum sensing by inhibiting LuxR activation, thereby disrupting bacterial communication and inhibiting biofilm formation (26) (27) In addition, naturally occurring compounds isolated from plants have shown inhibitory activities against quorum sensing. In particular, furocoumarins found in grapefruit juice are recognized as compounds that block both the AI-1 and AI-2 signal pathways, leading to significant biofilm inhibition in Salmonella typhimurium, Pseudomonas aeruginosa, and Escherichia coli O157:H7. These observations imply that compounds isolated from plants, such as furocoumarins, have the potential to act as natural antibiofilm compounds by interfering with the quorum sensing processes of microorganisms.(28)
MATRIX DEGRADING ENZYM
Biofilm matrix, which is dominated by polysaccharides, proteins, and extracellular DNA (eDNA), confers structural reinforcement and antimicrobial resistance. Enzymes like Dispersin B and DNase I can break down biofilm structures, inhibit adhesion, and increase the effectiveness of antimicrobial agents like povidone-iodine.(6)
In addition, α-amylase, a polysaccharide-breaking enzyme, has been found to interfere with Staphylococcus aureus biofilms. α-amylase commercial products will prevent cell aggregation, remove preformed biofilms, and inhibit biofilm formation.(29)
INHIBITION OF BACTERIAL CELL DIVISION
Certain antimicrobial peptides, such as proline-rich peptides such as pyrrhocoricin, disrupt intracellular protein folding by inhibiting the chaperone protein DnaK from carrying out its function. DnaK aids in bacterial stress response and protein repair. Pyrrhocoricin, by inhibiting DnaK from acting, inhibits vital bacterial activities and reduces biofilm formation and maintenance.(30)
CLEAVAGE OF PEPTIDOGLYCAN
Disruption of peptidoglycan, a structural molecule of the cell wall, may disrupt the biofilm and decrease bacterial adhesion. Natural products such as tannic acid may induce IsaA, S. aureus lytic transglycosylase, that hydrolyses the β-1,4 glycosidic bond between MurNAc and GlcNAc, disrupting the cell wall.(31) (32) Hydrolysis may also liberate biofilm-controlling signalling molecules and affect anchoring surface proteins.
PREVENTION VIA BIOFILM DISASSEMBLY
During biofilm disassembly, embedded cells are released and the extracellular matrix is actively broken down. Proteases, DNases, and surfactants can all initiate this process, which is frequently controlled by quorum-sensing systems like Staphylococcus aureus's agr system.(33) Another way to cause biofilm breakdown is to target amyloid-like fibres, including curli in E. coli and TasA in Bacillus subtilis. Parthenolide and FN075 are examples of inhibitors that disrupt these structural proteins. Likewise, D-amino acids, such as D-tyrosine, can cause disintegration at low levels, although in vivo toxicity is still an issue.(34) Usnic acid is one example of a natural substance that has antibiofilm activity by decreasing EPS synthesis and disrupting quorum sensing pathways, which results in changed biofilm shape and decreased biofilm bulk.(35) (36) In brief antibiofilm agents and their mechanisms is provided in Table 1.
Combining antibiofilm agents with conventional antibiotics significantly enhances therapeutic outcomes by disrupting the biofilm barrier and increasing drug penetration. These strategies represent a promising direction in the management of chronic and device-associated infections.

Figure 3: Schematic representation of key mechanisms by which agents disrupt biofilms. These include inhibition of quorum sensing, degradation of the extracellular matrix, disruption of cell division, cleavage of peptidoglycan, biofilm disassembly, and proteolytic activity.
TABLE 2: OVERVIEW OF ANTIBIOFILM AGENTS AND THEIR MECHANISMS OF ACTION
|
Category |
Agent /Example |
Mechanism of Action |
Target Organisms |
Notes |
|
Quorum Sensing Inhibitors |
Synthetic AHL analogs |
Block LuxR activation to disrupt quorum sensing |
P. aeruginosa, E. coli |
Inhibits biofilm gene regulation |
|
|
Furocoumarins (grapefruit juice) |
Inhibit AI-1 and AI-2 signaling pathways |
S. typhimurium, P. aeruginosa, E. coli |
Plant-derived, natural quorum sensing blocker |
|
Matrix-Degrading Enzymes |
Dispersin B, DNase I |
Degrade eDNA and PNAG to disrupt matrix and enhance antimicrobial action |
S. aureus, S. epidermidis |
Used in conjunction with iodine or antibiotics |
|
|
α-Amylase |
Breaks down extracellular polysaccharides |
S. aureus |
Reduces preformed biofilms and inhibits aggregation |
|
Cell Division Inhibitors |
Pyrrhocoricin |
Inhibits DnaK chaperone protein, interfering with protein folding and stress response. |
Broad spectrum (G+) |
Reduces bacterial viability and biofilm sustainability |
|
Peptidoglycan Cleaving Agents |
Tannic Acid |
Upregulates IsaA, a lytic transglycosylase, weakening cell wall |
S. aureus |
Does not harm planktonic viability |
|
Biofilm Disassembly Agents |
Proteases, DNases, surfactants |
Degrade matrix and promote detachment |
S. aureus, B. subtilis, E. coli |
Often regulated by agr and other QS systems |
|
|
Parthenolide, FN075 |
Inhibit amyloid fiber (TasA/curli) formation |
B. subtilis, E. coli |
Target protein–protein interactions in matrix assembly |
|
|
D-Tyrosine, D-Cysteine |
Alter EPS production; disrupt biofilm architecture |
P. aeruginosa, A. baumannii, B. subtilis |
Effective in vitro; toxic in vivo at high concentrations |
|
|
Usnic Acid |
Inhibits EPS production, quorum sensing, and hyphal transition |
S. aureus, Candida albicans, P. aeruginosa |
Lichen-derived; broad-spectrum antibiofilm activity |
BROMELAIN
Bromelain is a cysteine protease enzyme complex first identified in Brazilian pineapple by Peckold et al. (37) is primarily extracted from the stem and fruit of the pineapple plant (Ananas comosus Merr.), with the stem being the major commercial source other parts such as the pineapple core and pulp also offer cost-effective alternatives for bromelain production.(38) The enzymatic activity of fruit bromelain and stem bromelain varies depending on the extraction source, with stem bromelain showing greater enzymatic potency at least eight different isoforms of bromelain have been identified from the stem; each is made up of a heavy chain with five disulfide bonds and a light chain. A sulfhydryl proteolytic enzyme that is unique to cysteine proteinases ) is the main active ingredient (39) (40)
Crude bromelain is a complex enzymatic mixture, that including cysteine proteases like fruit bromelain, stem bromelain, ananain, and comosain as well as other enzymes such as phosphatases, peroxidases, ribonucleases, cellulases, protease inhibitors, glycoproteins, carbohydrates, and organically bound calcium.(41) This diverse biochemical composition gives bromelain's potent antimicrobial, anti-inflammatory, and wound-healing properties, makes it as a promising agent in biomedical and wound care applications.
Bromelain exhibits optimal activity around pH 6–7 and has an isoelectric point (pI) of 9.55(42) glass transition temperature is about 61 °C, as determined by Differential Scanning Calorimetry (DSC), indicates that drying should be conducted below this threshold to preserve activity. (43) While its simple extracts show peak activity at pH 7 and 50 °C, ethanol- precipitated forms optimal performance at pH 8 and 60 °C. (44) Bromelain has shown promise as an oxidizing agent in addition to its proteolytic activity.(45) Crucially, bromelain has been used extensively as a bioenhancer for antibiotics because of its low systemic cytotoxicity, which has greatly improved antimicrobial outcomes in diseases like sinusitis, pneumonia, bronchitis, and skin infections brought on by Staphylococcus aureus. (46) (47) According to a pharmaco-kinetic studies by bromelain taken orally can be absorbed in trace amounts in a physiologically active, with detectable plasma concentrations that last for up to 48 hours. This reflects its potential for systemic treatment when taken orally for wound or infection treatments.(48)
ANTIBIOFILM AND ANTIBACTERIAL ACTIVITY
Bromelain is a natural choice for the treatment of periodontal infections because it has demonstrated antibacterial action against important dental and skin pathogens such S. mutans and P. gingivalis.(49) (50) bromelain not only showed intrinsic antibacterial action against Staphylococcus aureus and Staphylococcus epidermidis, but it also improved the effectiveness of 14 other antibiotics, particularly those that target Gram-negative bacteria. Interestingly, amoxicillin, erythromycin, ciprofloxacin, and gentamycin all showed synergistic benefits. (51)
As antibiofilm agent bromelain, particularly in combination with N-acetylcysteine (BromAc), has shown remarkable potential. In vitro studied have demonstrated that BromAc dissolve more than 80% biofilm dissolution in Pseudomonas aeruginosa, underscoring its potential utility in managing chronic and device-associated infections. (11) furthermore 1% bromelain significantly inhibited biofilm formation (up to 4-fold) and effectively disrupted mature biofilms (up to 6.4-fold), with the strongest effects observed in methicillin-resistant S. aureus strains. These effects are attributed primarily to its proteolytic activity, with some support from DNase-like action.(52)
WOUND HEALING AND ENZYMATIC DEBRIDEMENT
Bromelain aids in the efficient removal of tissue in chronic wound models and has been used successfully in enzymatic wound debridement. Clinical trials highlight its usefulness as a safe, non-surgical alternative for wound treatment by reporting an average debridement of 68% with no significant side effects observed. (53) Bromelain-loaded nanoparticles (NPs) improved re- epithelialization in animal wound models by enhancing sustained enzymatic release when added to chitosan-based hydrogels.(54)
TABLE 3: KEY STUDIES ON BROMELAIN’S ANTIBACTERIAL, ANTIBIOFILM, AND WOUND-HEALING ACTIONS
|
Study |
Target Organism |
Key Findings |
Mechanism |
Notes |
|
Praveen et al., 2014 |
Streptococcus mutans, Enterococcus faecalis, Aggregatibacter actinomycetemcomitans, Porphyromonas gingivalis |
Bromelain showed antibacterial activity with MICs ranging from 2–31.25 mg/mL |
Proteolytic action; antibacterial |
Most effective against S. mutans (2 mg/mL) and P. gingivalis (4.15 mg/mL); indicates potential as adjunct in periodontal therapy |
|
Carter et al., 2021(11) |
Pseudomonas aeruginosa (3 strains) |
Bromelain + NAC (BromAc) dissolved >80% biofilm on mesh in vitro |
Synergistic mucolytic activity |
Strain-dependent; NAC alone promoted growth in 2 strains |
|
Bayat et al., 2022(55) |
Wound models (animal) |
Bromelain NPs in chitosan hydrogel enhanced debridement and re-epithelialization |
Sustained enzymatic release |
92% encapsulation; stable at 4 and 25 °C |
|
Shoham et al., 2021(56) |
Chronic wounds (clinical, various etiologies) |
68% average debridement; effective in venous/diabetic wounds |
Topical enzymatic proteolysis |
Safe, non-surgical alternative; mild pain only adverse event |
|
Silva et al., 2023 (57) |
Staphylococcus aureus (incl. MRSA) |
1% bromelain inhibited biofilm formation (4×) and disrupted biofilms (6.4×) |
Proteolytic activity; some DNase effect |
No MIC effect; strong action on MRSA from chronic wounds |
|
Omotoyinbo et al., 2025 |
Staphylococcus aureus, Staphylococcus epidermidis, various Gram-negative bacteria |
Bromelain enhanced antibiotic efficacy and showed inherent antibacterial activity; optimum activity at 40°C, pH 7.0 |
Synergistic antibacterial action; improved antibiotic absorption |
Crude and purified bromelain tested; best synergy observed with β-lactam and macrolide antibiotics |
COMPARATIVE EVALUATION OF BROMELAIN AND OTHER NATURAL ANTIBIOFILM AGENTS
Numerous natural substances with different modes of action have demonstrated anti-biofilm qualities. Because of its combined potential to increase antibiotic permeability and break down the components of the biofilm matrix, bromelain stands out from the others. More research is being done on the possibility of bromelain as a therapy, both on its own and in conjunction with other natural anti-biofilm medications.
The antibiofilm and antibacterial activity of flogomicina, a natural antioxidant mixture of NAC, bromelain, and other plant extracts, was significantly stronger than that of NAC alone. It reduced E. Coli and P. mirabilis growth by over 80% and improved amoxicillin efficacy over 14 days, indicating a synergistic effect and the potential for reducing antibiotic dosage.(58) Manuka honey, tea tree oil, and phytochemicals such as usinc acid, tannic acid, and eugenol also exhibit significant antibiofilm activity. (59) (60)
Tea tree oil (TTO) and its active ingredient terpinen-4-ol have been shown to have effective antibiofilm activity against Candida albicans, even resistant strains. A 60-second exposure to either TTO (17.92 mg/mL) or terpinen-4-ol (8.86 mg/mL) significantly inhibited biofilm growth, suggesting their potential use as natural antifungal agents in the treatment of oral candidiasis.(61) Tannic acid inhibits biofilm formation of E coli by targeting the curli subunit gene D (CsgD) regulators, which controls matrix development and curli production, additionally it also modulate biofilm regulated gene expression and enhance signalling of indole, highlighting its potential as antibiofilm agent.(62)
A lichen derived compound usnic caid, shown strong antibiofilm activity by preventing adhesion, EPS production and virulence factors in Pseudomonas aeruginosa, Staphylococcus aureus. It makes promising natural antibiofilm candidate by biofilm structure and reduce biomass.(59) α-Amylase is a glycoside hydrolase, it effectively disrupts Pseudomonas aeruginosa, Staphylococcus aureus biofilms in both monoculture and mixed species model. These enzymes degraded the extracellular matrix, reduced biofilm biomass, and enhanced the efficacy of antibiotics in vitro and in vivo. According to the study, glycoside hydrolase treatment is a safe and effective way to treat chronic wound infections linked to biofilms.(63)
Consequently, bromelain can be used as standalone antibiofilm agent and also as combination therapies for synergetic effect. Further comparative analysis is needed to find mechanistic diversity, relative efficacy and clinical potential of bromelain versus natural other antibiofilm agents in both mono and polymicrobial wound infections.
TABLE 4: COMPARATIVE SUMMARY OF NATURAL AND ENZYMATIC ANTIBIOFILM AGENTS
|
Agent |
Source/ Type |
Mechanism of Action |
Target Organisms |
|
Bromelain |
Enzyme obtained from pineapple stem |
Biofilm matrix proteolysis, enhance antibiotic penetration. |
S. aureus, P. aeruginosa, E. coli, MSRA |
|
Tannic Acid |
Polyphenol (plants) |
Represses CsgD regulator, synthesis of curli, and biofilm- related genes. |
S. aureus, Agrobacterium tumefaciens, E. coli, |
|
Usnic Acid |
Lichen derived compound |
interferes with virulence genes, adhesion, and EPS structure. |
S. aureus, P. aeruginosa, Candida albicans |
|
Tea Tree Oil |
Essential oil (Melaleuca) |
breaks down membranes; terpinen-4-ol prevents metabolism and adhesion. |
C. albicans, S. aureus, E. coli |
|
α-Amylase |
Enzyme (microbial/plant) |
EPS's extracellular polysaccharides are hydrolysed. |
S. aureus, P. aeruginosa, MRSA, V. cholerae |
CONCLUSION
The presence of robust microbial biofilms that delay healing and contribute to antibiotic resistance makes chronic wounds persistent therapeutic issue. These bacterial communities, which are dominated by pathogens like Pseudomonas aeruginosa and Staphylococcus aureus, use complex defence strategies include altered cell physiology, extracellular matrix formation, and quorum sensing. Innovative approaches to therapy must be explored because conventional treatments often prove insufficient against these well-established biofilms. Natural agents show promising antibiofilm agents due to different mechanism of action, synergistic activity with conventional therapies and biocompatibility. This review reflects several affective strategies; quorum sensing inhibition, enzymatic matrix degradation, disruption of cell division, and biofilm disassembly. Among these approaches bromelain exhibits exceptional versatility through proteolytic activity, synergistic effect with antibiotics, biofilm disruption capability, wound healing enhancement. The clinical evidence that support bromelain's efficacy includes significant biofilm inhibition (up to 4-fold reduction in formation, 6.4-fold reduction in mature biofilms), enhanced antibiotic synergy.
FUTURE PROSPECTS
Future research should focus on optimized delivery system with standardized formulations, and well-designed clinical trial for comparing bromelain-based therapies with existing treatment for diverse wound type. Furthermore, combination therapy demonstrating bromelain synergistic effect with conventional antibiotics and with other natural agents to maximize therapeutic outcome while minimizing adverse effects. Integrating cost effective analysis, patient reported outcome, long term follow-up in to future trials will provide correct understanding of bromelain effect in wound treatment. This direction will help for robust evidence-based procedure for broader clinical application.
ACKNOWLEDGMENT
The author expresses sincere gratitude to all researchers whose work has been cited in this review. Appreciation is also extended to those who provided insightful feedback and moral support during the preparation of this article. Their contributions were invaluable to the completion of this work.
REFERENCES





Twinkle N. U., Shijith K. V., Ansira P., Arya P. V., Baby Nishma, Targeting Wound Biofilms: Biofilm Formation, Resistance, and Natural Antibiofilm Strategies, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 7, 3136-3150. https://doi.org/10.5281/zenodo.16356613
 10.5281/zenodo.16356613
10.5281/zenodo.16356613
