Department of Pharmacy, Institute of Technology and Management, GIDA, Gorakhpur, Uttar Pradesh, India, 273209 Affiliated to- Dr. A.P.J. Abdul Kalam Technical University, Lucknow, Uttar Pradesh, India
Triple-negative breast cancer (TNBC) is an aggressive subtype of breast cancer lacking estrogen receptors, progesterone receptors, and HER2 expression, which limits targeted therapeutic options and contributes to poor clinical outcomes. D-?-Tocopheryl polyethylene glycol 1000 succinate (TPGS), a water-soluble derivative of natural vitamin E, has emerged as a promising nanocarrier due to its amphiphilic nature, ability to enhance drug solubility, inhibit P-glycoprotein (P-gp), and selectively induce apoptosis in cancer cells. This review explores the physicochemical and pharmacological properties of TPGS and its application as a nanocarrier in TNBC therapy. TPGS induces cytotoxic effects via multiple mechanisms, including P-gp inhibition, reactive oxygen species (ROS) generation, mitochondrial dysfunction, and DNA damage, leading to caspase-dependent and independent cell death. It also enhances the therapeutic efficacy of chemotherapeutic agents by overcoming multidrug resistance. Furthermore, the paper discusses the molecular heterogeneity and complex signaling pathways involved in TNBC pathogenesis—such as PI3K/AKT/mTOR, Notch, Wnt/?-catenin, and Hedgehog—which offer potential targets for combinatory or synergistic treatment strategies involving TPGS-based nanoformulations. These findings support the role of TPGS as a multifunctional excipient and therapeutic agent in addressing the challenges associated with TNBC treatment.
Cancer is the main reason of death for individuals under 70 in 57 out of 183 countries, including the majority of Europe and North America, and the second leading cause of death worldwide. Breast cancer being the sixth leading cause of cancer related mortality, and it is the most common female malignancy worldwide other than non-melanoma skin cancer [1. It is estimated that by 2040, there would be one million yearly fatalities and over 3 million new cases [2]. HER-2 (human epidermal growth receptor 2), progesterone receptor (PR) and estrogen receptor(ER) expression classes are to identify the types of breast cancer molecular subtypes. Breast cancer is neither simple disease. Luminal A: ER and PR positive, HER-2 negative; luminal B: The ER is positive, PR-low/ negative HER-2-negative in HER-2 positive breast cancer where HER-2 is overexpressed and/or her-2 is amplified, ER and PR are negative Meanwhile, Triple-negative breast cancer (TNBC) lacks the above 3 receptors [3]. TNBC, the most aggressive subtype of breast cancer is associated with higher risk for distant relapse and death within 5 years of diagnosis [4]. Approximately 46% of TNBC patients are predicted to develop metastases in several organs, including as the liver, bone, central nervous system, and lung/pleura [5]. The lack of specific cell receptors in TNBC presently limits the available treatment options. Despite being the mainstay of the treatment plan, chemotherapy is not very successful when used as a postoperative adjuvant [6]. Even with early therapy, most TNBC patients still have residual disease, which substantially reduces survival in the first three to five years and increases the likelihood of relapse [7]. The amphiphilic macromolecule is D-alpha-tocopheryl polyethylene glycol 1000 succinate (TPGS), a water-soluble natural vitamin E derivative. It works well as a nanotechnology emulsifier in biomedicine. In addition to improving solubility and cellular uptake, TPGS co-administration can boost the oral bioavailability of a variety of anticancer drugs and block the P-glycoprotein-mediated multi-drug efflux transport mechanism. By esterifying vitamin E succinate with polyethylene glycol (PEG) 1000, vitamin E TPGS—a water-soluble version of natural vitamin E—is created [8]. PEG is the most often used hydrophilic segment because of its remarkable biocompatibility and water solubility. In order to form the molecule so that its molecular weight is larger than that of the hydrophobic core, the micellar shell is usually chosen. Critical micellar concentration is generally in the micro molar range of these findings in micelles and less than 100 nm; Vitamin E TPGS —Nonionic surfactant with MW rate of 1513 g.mol-1 and polar head amphiphilic frame fully soluble in water, containing lipophilic alkyl tail and hydrophilic polar head amphophilic backbone. It stays stable and degrades less than 12 percent in the pH range of 4.6 to 7.6 after three months in a neutral aqueous buffer. It has been found that the oral LD50 for vitamin E TPGS safety in adult male rats is more than 7,000 mg/kg [9–10]. Moreover, a variety of molecules, including as hormones, antibiotics, taxans, cyclosporine, and both water-soluble and water-insoluble compounds, can be dissolved by vitamin E TPGS [10]. It's possible that vitamin E TPGS inhibits P-gp. It is more effective in preventing P-gp from working than other non-ionic surfactants like Tween 80, Pluronics, and Cremophor. It has been used in several formulations and applications, including as the production of nano suspensions [11]. Adjuvants for vaccination systems, nanoparticles, dependent prodrugs, potent dispersions/tablets, and self-micro emulsifying nutrition supplements [12–15]. Vitamin E TPGS is a hydrophobic drug carrier that enhances eye permeability and promotes ocular retention, among its many other applications. TPGS is used to treat vitamin E deficiency or as a vitamin E supplement when a person is unable to consume fats due to a specific ailment [16].
An overview of vitamin E and TPGS
Tocopherols (TOS) and tocotrienols are two forms of vitamin E, which is a lipophilic material. Human tissues and plasma contain a large amount of the vitamin α-TOS. This type controls the anti-inflammatory and antioxidant characteristics of cells by inhibiting protein lipoxygenase enzymes. In addition, α-TOS plays a pivotal role in safeguarding the RBC membrane and lung cells from free radicals toxicity [16]. Several α-TOS analogues with increased biocompatibility and anticancer efficacy were readily obtained through chemical modification of the chromanol (head group) and phytyl (tail group) modifications of vitamin E. An example of the modification is TPGS [α-TOS-succinyl-Poly (1–2-oxatride)} iso]ón. PEG, which has chemical formula C33O5H54 (CH2CH2O) n, is used to esterify α-TOS succinate. The number "n" indicates how many PEG molecules are joined. TPGS is amphiphilic due to its composition of hydrophilic PEG.and lipophilic α-TOS TPGS has been shown to be safe and non-genotoxic. In many nano formulations developed for tumor therapy, it is the preferred excipient (adjuvant) due to its strong anticancer potential as compared to plain α-TOS.
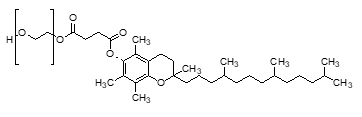
Chemical structure of TPGS
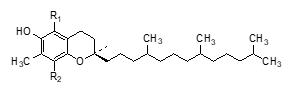
Chemical structure of Vitamin E
Classification of triple-negative breast cancer (TNBC)
Tumor Pathogenesis of Triple Negative Breast Cancer
The toughest category of breast cancer known as TNBC. No HER2 receptor amplification, PRs or ERs. TNBC consists of both negative ER and PR in lateral myoepithelial cells of breast lobules as well as negative HER2 expression on the epithelial cells of breast. Missed, less than 1% ER or PR detect out of staining by immunohistochemistry + HER2 score of 0/+1 or +2 with FISH negative) [17-18]. ER, PR and HER2 negative cells are the most vulnerable type of this mutation in BRCA gene Another DNA damage checkpoint and component of DNA repair, BRCA1. It turns ER-negative cells to ER-positive and is required for mammary gland development of course ER-negative only. BRCA1 and BRCA2 fix the double-stranded break in DNA using homologous recombination. The homologous recombination is controlled by ATM, BRCA1 and BRCA2. DNA damage cause the ATM to be phosphorylated which then initiates the homologous recombination process in association with RAD51 and BRCA2 [19]. Genomic instability is the outcome of the inability for the BRCA gene mutation to be repaired by homologous recombination because it enables mutagenesis processes to fix DNA double strand breaks. The origin of Cancer arising from this genetic instability. Another factor that is part of the node of TNBC is alterations in the activity of mTOR pathway of mammal. This change leads the disease to become aggressive and invasive. Activation of the PI3K/AkT/mTOR (PAM) cascade in TNBC drove cancer cells to proliferate, grow and angiogenize. When mTOR is activated, two further mTOR complexes are created i.e mTORC1 and mTORC2. mTORC1 stimulates cell proliferation, whilst mTORC2 is needed to phosphorylate AKT [20-21]. The tumor cells have poor control over cell cycle progression, they either allow genesis of genetic diversity or license cells with DNA pathology to proceed to next cell in the cycle. TNBC cells are frequently characterized by genetic alterations of tumor suppressor protein (p53) and Rb pathway inactivation rather than c-Myc pathway mutations [ 15 ]. Phosphorylation by cyclin/Cdk complex activation of retinoblastoma initiates the Rb pathway at G1/S checkpoint of cell cycle control: which subsequently phosphorylates transcription factor E2F by Myc. Conversely, up regulation of CDNK2 by makes CDK2 less active inactivate the suppressor gene p53 that prevetn senescence in apoptosis pathways.Increased expression of CDNK2A and the related CDNK20 complex allows for the G1/S phase to be reached and the cell-cycle regulatory checks in the Rb pathways to be avoided. As a result, it causes cellular development and proliferation to be expressed more. TNBC is brought on by this recurring genetic change [22–23]. Figure 1 depicts the typical Triple negative breast cancer pathways.
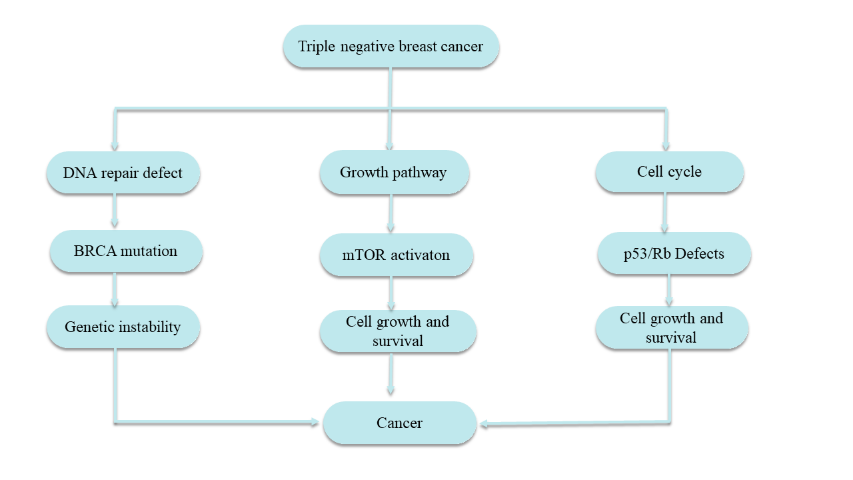
Figure 1. Triple negative breast cancer pathways
Mechanism of action of TPGS on triple negative breast cancer
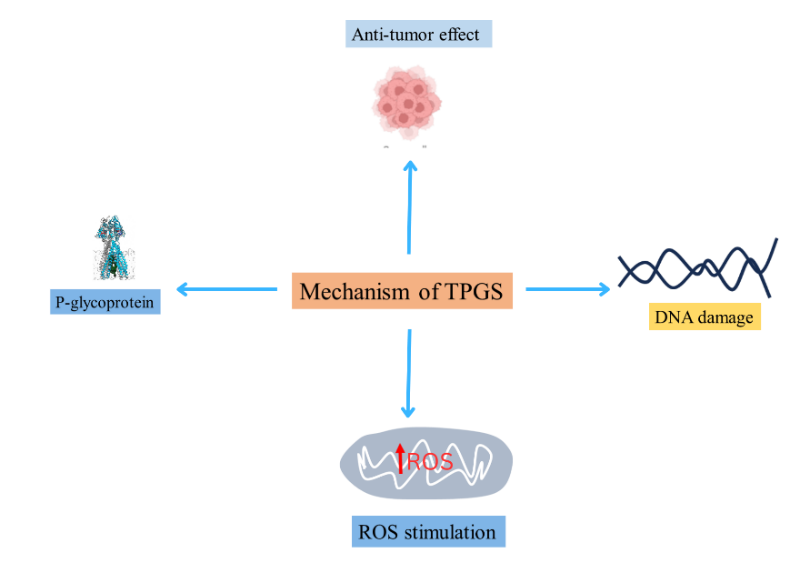
Figure 2. The effects of TPGS, including P-glycoprotein inhibition, ROS stimulation, DNA damage, and anti-tumor activity.
TPGS has anticancer properties. TPGS can cause apoptosis and exhibit selective cytotoxic activity against in vitro cancer cells through synergistic antitumor action. These effects can be mitigated and therapeutic efficacy increased by loading or combining TPGS with chemotherapeutic medicines. Following TPGS treatment, the reaction of cancer cells and healthy immortalized breast cells is highly valuable. In breast cancer cell culture, TPGS can cause G1/S cell cycle arrest by triggering the apoptotic signaling pathways [24].
Eliminations and suppressions with respect to mitochondrial respiratory complex II [25] that would kill apoptosizing cancer cells ( alpha-tocopheryl succinate alpha-TOS) The secondarily electroned defective transport chain could cause ROS. The upregulation of intracellular ROS, a mediator in DNA damage and protein/ lipid/enzyme oxidation leading to apoptosis [26], Is. Given that tumor cells may be more prone to ROS, this is also in part the rationale behind selective cancer cell killing by anticancer drugs. Antiapoptotic proteins, such as the downregulation of anti-apoptotic protein Bcl-2 that can activate caspase-3 and caspase-7 (caspase-dependent programmed cell death) as well as Surviving, which forgoes the action of apoptosis e. from being inhibited after phosphorylation of protein kinase B [27], might be downregulated by TPGS. Also, being at the same time a G1/S phase cell cycle arrest and caspase-independent programmed cell death. Mitochondrial associated apoptosis and ROS production, TPGS generally also seems to be detrimental to cancerous cells, including breast cancer and lung adenocarcinoma [28].
Both caspase-independent and caspase-dependent DNA damage can be induced by TPGS [29]. Since vitamin E can prevent tumor resistance often linked to the use of traditional chemotherapeutic medicines that cause caspase-dependent programmed cell death, its capacity to induce caspase-independent programmed cell death may in fact be useful in breast cancer treatment. [30].
The P-glycoprotein (P-gp or MDR protein 1; MDR-1) is an ATP dependent drug efflux pump on the cell membrane, leading to the low accumulation of lipophilic drugs in P-glycoprotein expressing Multidrug Resistance (MDR) cell lines.
It influences drug absorption and distribution, metabolism as well as excretion in the intestinal epithelium, blood-brain barrier (BBB), blood -testis barriers, hepatocytes, renal proximal tubular epithelial cell and adrenal cortex as well as in capillary endothelial cells [31]. P-gp overexpression within cancer cells would also compromise drug entry and therefore the antitumor activity arising from such excessive accumulation medications. Consequently the most successful way to lessen MDR is to use an inhibitor of P-gp. Beneath TPGS concentration in CMC (0.02 wt%) it was demonstrated that P-gp activity is dysfunctional [15]. Other P-gp inhibitors, such as Tween 80 and Cremophor EL (nonionic surfactants) were avoided due to deleterious effects.
Pharmaceutical properties of TPGS
TPGS1000 is a water-solubility form of vitamin E100. Although water miscible, it can be also supplemented with co-solvents such as propylene and polyethylene glycols, oils, as well as other surfactants in its composition. Agenerase®, same as many commercial products based on TPGS being used as solubilizer. TPGS has been a utilized formulation of an excipient used in nanoformulation for absorption enhancer and solubilization too. Flor et al., for instance, intended to improve the solubilization and bioavailability of NFV by employing new TPGS1000 succinate micelles, which are carrying NFV mesylate. They discovered that NFV's solubility increased by 18 times in micelles made with TPGS as a solubilizer [32].
Vitamin E TPGS is used as a source of water-soluble vitamin E and as a permeation enhancer in beauty products like lotions and creams, as well as personal care products like eye drops and nasal sprays. It functions as a hypoallergenic, ethanol-free, non-irritating emulsifier/excipient in cosmetic and personal care goods [33].
In nano particulate systems, TPGS has been shown to be an excellent emulsifier and stabilizer. Compared to the traditional polyvinyl alcohol (PVA) emulsifier, it has demonstrated a 67-fold higher emulsification efficiency. Paclitaxel NPs emulsified with Vitamin-E TPGS were synthesized by Zhao and colleagues. Compared to other emulsifiers like PVA, they found that TPGS had superior biocompatibility and emulsification properties. Additionally, NPs created with TPGS as an emulsifying agent demonstrated stability, enhanced cellular absorption of NPs by cancer cells, excellent cargo encapsulation efficiency, and sustainable pharmacokinetics [34].
When creating films such as PLLA (poly lactic acid) and HPC (hydroxyl propyl cellulose), among others, TPGS can be used as a plasticizer. According to tensile testing, it increases the films' flexibility and elongation at the flouting point while lowering the glass transition temperature and adhesive force. Because TPGS creates a unique structural film, it can be used to prepare various implants intended for localized drug delivery. It has been shown to increase the biocompatibility of poly sulfone hollow fiber membranes for hemodialysis in blood purification [35].
Heterogeneity of TNBC
Heterogeneity refers to the phenomena in which various cancer cells have varied cellular morphologies, gene profile, proliferative capability, and metastasis. TNBC displays heterogeneity both within and between tumors [36]. Heterogeneity arises secondary to disparate somatic mutations in the genes for TNBC as well. To further dissect the heterogeneity of TNBC, the TCGA Research Network explored early breast cancer cells using six platforms: DNA methylation, exome sequencing, microRNA sequencing messenger RNA arrays,Copy number platforms as well as reverse-phase protein arrays. Combining data from the six platforms, TCGA indicated that the heterogeneity in TNBC is predominantly caused by alterations in DNA damage repair genes. This includes impairment of the functions of BRCA1, retinoblastoma protein 1 (RB1), and tumor protein 53 (TP53) [37]. Furthermore, TCGA observed mutations in the signaling pathways of phosphoinositide 3-kinases (PI3K) and mammalian target of rapamycin (mTOR) as a result of the loss of lipid phosphatases INPP4B (inositol polyphosphate 4-phosphatase type II) or PTEN (phosphatase and Tensin homolog). But when it was noted that certain individuals exhibited less mutation at diagnosis while others displayed fast mutation, heterogeneity became more supported [38]. It is possible to conclude that chemotherapy has a distinct effect on various parts of a single tumor as a result of this mutation in triple negative breast cancer cells. It is also possible that the heterogeneity will make TNBC cells resistant to treatment. [37]
Potential Therapeutic Targets for TNBC Therapy
Main disadvantage of chemotherapy is the high TNBC heterogeneity. Furthermore, TNBC is considered an aggressive subtype of breast cancer in that it metastasizes to the brain and other visceral organs very fast and patients with advanced TNBC will generally die from their disease within an average of about 12 months the next round survive. Now that is a pretty big deal actually to be able to treat TNBC with specific targets. In the next part I would like to mention the possible targets of TNBC. Byway of example, they include angiogenesis, Hedgehog (Hh), Notch, the Wnt/-catenin signaling networks, PI3k/AKT/mTOR pathways; as well as other specific receptors such as EGFR, IGF 1R, PARP1 targeted receptors, programmed cell death ligand 1 pathway (PDL-1) targeted receptors,cytotoxic T lymphocyte antigen 4 targeted receptors (CTLA-4), chondroitin sulfate proteoglycan pathway 4 (CSPG4) protein targeted receptors and androgen receptors (AR).
Notch Signaling Pathways
The Notch signaling system regulates numerous essential cellular activities and is assumed to be an extremely conserved specialized mechanism that requires juxtracrine (cell-to-cell) interaction. [40]. It is well known that notch signaling controls differentiation and self-renewal, both of which are necessary for the mammary gland to mature normally [41–42]. The contact between the Notch ligand and the Notch receptor initiates the signaling cascade. Five Notch ligands and four receptors have been found, including Jagged (JAG)-1 and 2 and Delta-like (DII)-1, 3, and 4. Every Notch ligand is a transmembrane protein that has several EGF-like repeats and an extracellular DSL domain. The extracellular DSL domain mediates the binding process. Cysteine is abundant in an additional domain of the JAG ligand. [43]. The proteolytic enzymes ADAM and TACE help the Notch ligand attach to the receptor, releasing the receptor's ectodomain and forming a Notch ligand-receptor complex. [44–45]. The Notch ligand-receptor complex is subsequently changed into NEXT (Notch extracellular truncation) by the proteolytic action. The Notch intracellular domain (NICD) is released after the NEXT is finally broken down by secretase. It then moves from the cytoplasm to the nucleus [46] and binds with the transcriptional activator, specifically Mastermind (MAML1-3, and MED3) and the CSL complex (CBF1, RBPJK/Su(H)/LAG1) [47,48].
Angiogenesis and apoptosis modulators, cell cycle regulators, growth factor receptors, transcriptional factors like NF-B2 and c-Myc, and genes like ER, Hes, Hey, and VEGFR3 are among the downstream targets whose transcription is triggered by the binding, which activates the CSL-complex. [49] Dysregulation of the Notch signaling pathway has been reported to commonly result in aberrant 9 of 64 stem cell self-renewal and differentiation, causing carcinogenesis. [50-51]
Hedgehog (Hh) Signaling Pathway
Hh signaling is an evolutionarily conserved pathway that plays an important role during embryonic mammary gland morphogenesis, ductal morphogenesis and epithelial-mesenchymal transition (EMT) [52]. The Hh signaling is a pathway made up of three ligands Sonic Hedgehog (Shh) Desert Hedgehog (DHh) and Indian Hedgehog ( Ih ) In the most part, their target ligands are Sonic Hedgehog. The PTCH Receptor which suppresses co-receptor SMO, [3] functions indirectly through SMO co-receptor. That enables the assembly of the GLI complex, a feature of Hh activation. The GLI complex is dependent on the transcriptional factors GLI1, GLI2 and most prominently GLI3. It is during activation that GLI complex is targeted and translocates into the nucleus : this increases the expression of GLI1 & GLI2 transcription factors causing metastasis, angiogenesis and apoptosis ultimately contributing to TNBC development [57]. [53] also increases expression of metastasis proteins, (SNAIL) and angiogenesis [54-55] factors (angiopoietin-1, and 2). In addition, SMO is directly over-expressing whole MYCN, consequently incrementing the expression of transcription factors FOXM 1 [51] and cyclin D [56] in TNBC cells.
Wnt/-Catenin Pathway
Wnt signaling pathway, important embryonic tissue patterning; cell migration, adhesion; stem cell self-renewal and epithelial mesenchymal interactions. Wnt binding to LDL receptor-related protein 5/6 together with Sfruzel (FZD; an 7-pass transmembrane protein) la:signal pathway activates-,Frizzled protein [57]• Lyn Wnt/ LRP5/6-Frizzled complex in the absence of Wnt pro teins/ ligands conceal-catenin?dehalved by a complex,involve axin, adenomatous polyposis coli (APC) tumor suppressor, glycogen synthase kinase-3 (GSK3), and casein kinase 1 (CK1), and then cause phosphorylation of-catenin by CK1 and GSK3 eventually leading to ubiquitination [23]. Then the ubiquitination causes degradation of-catenin by 26S proteasome [58]. Wnt proteins bind to LRP5/6 and FZD to form a Wnt-LRP5/6-FZD complex which in turn, prevent GSK3 and stabilize cytoplasmic-catenin Free -catenin then enters the nucleus, binds to T-cell factor/lymphoid enhancing factor and up-regulates targeted gene programs of cell growth &proliferation apoptosis through activation of TNBC [59-60]. Another study has highlighted up-regulated expression of FZD and LRP5/6 in TNBC cells as well [7]. In addition, the transcriptional loss of FZD/LRP6 by TNBC cells creates a hypothesis that it is held in check in vivo. It was generalized later that the association between the Wnt/-catenin pathway and TNBC advancement is associated with a gain in nuclear catenin or a loss of membranous-catenin [61]
TGF-Signaling Pathway
The growth, differentiation, homeostasis, and death of cells are all influenced by the TGF-signaling pathway. There are other members of the TGF-cytokine family, but TGF-1, which is encoded by the TGF-1 gene [62–63], has been shown to be crucial for breast cancer stem cells (BCSC). TGF receptor 1 (TGFBR1) overexpression has been noted in BCSC [64]. TGF-1 causes EMT in mammary cells, which leads to the development of tumors, according to another research [65]. A study by Sendurai et al. (2008) provided more support for this, showing that the mammary stem cells expressed more TGF-1, which enhanced their capacity to form mammospheres. Additionally, the cells expressed EMT genes such as N- and E cadherin, Slug, and Snail, which are linked to the progression of TNBC. According to Michael et al. (2011), cancer stem cells (SCs) produced by TGF-/TNF-induced EMT exhibited improved self-renewing ability in addition to higher tumorigenicity and resistance to chemotherapy [66]. Additionally, it was discovered that the TGF-1 pathway produces the production of SMAD2/3 and SMAD4, which results in angiogenesis, cell proliferation, metastasis, and protein synthesis. Therefore, it may be concluded that TGF signaling is essential for EMT activation and stemness acquisition; as a result, this route has been proposed as a possible treatment strategy against TNBCs.
PI3K/AKT/mTOR Signaling Pathway
PI3Ks is believed be one of the central molecules in PI3K/AKT/mTOR signaling cascade that fuel the tumorigenic potential of cancer cells. PI3Ks are primarily composed of heterodimers that are made up two whatsoever. However among PFIs release afull four PI3Ks isoforms as of day today that figuringpossess[67]. Receptors of tyrosine kinases give rise to the PI3K activatein KINASE» cascade that then PI3K activation — which on opposition on PI3K key was directerm [68], AKT phosphorylation [69], and mTOR. mTOR, the ser/thr kinase that regulates transcription, protein synthesis and cellular growth, motility and sustenance [69–70]. mTOR consists of any 2 complexes: mTORC1 and mTORC2, S-phase kinase association protein was activated through the both mTORC1 and mTORC2 in order that protein synthesis, cell growth or proliferation, angiogenesis as well metastasis occur. This has been identified that dysregulation of the mTOR pathway is linked to the growth of TNBC. Furthermore, besides overexpression of EGFR (upstream regulators) by PI3K/AKT/mTOR signaling pathway was predominantly through a study in TNBC breast cancers (71). [71]—at last, the hypotheses revealed tumor let and byfunction PIK3C mutational ptens expressionAL ow reduction due to the fact overexpression in the PI3K/AKT/mTOR signaling signaling pathway[71]
Treatment Strategies for Triple Negative Breast Cancer
TNBC is a heterogeneous aggressive form of cancer for which no scientifically validated biologically targeted effective treatment is approved yet. Fewer ERs, PRs, and HER2 means a very hard job of finding a stable alternative treatment for TNBC. TNBC will be discussed, in the next sections, a few of the most efficacious and robustly employed treatment strategies for TNBC and why they failed [72].
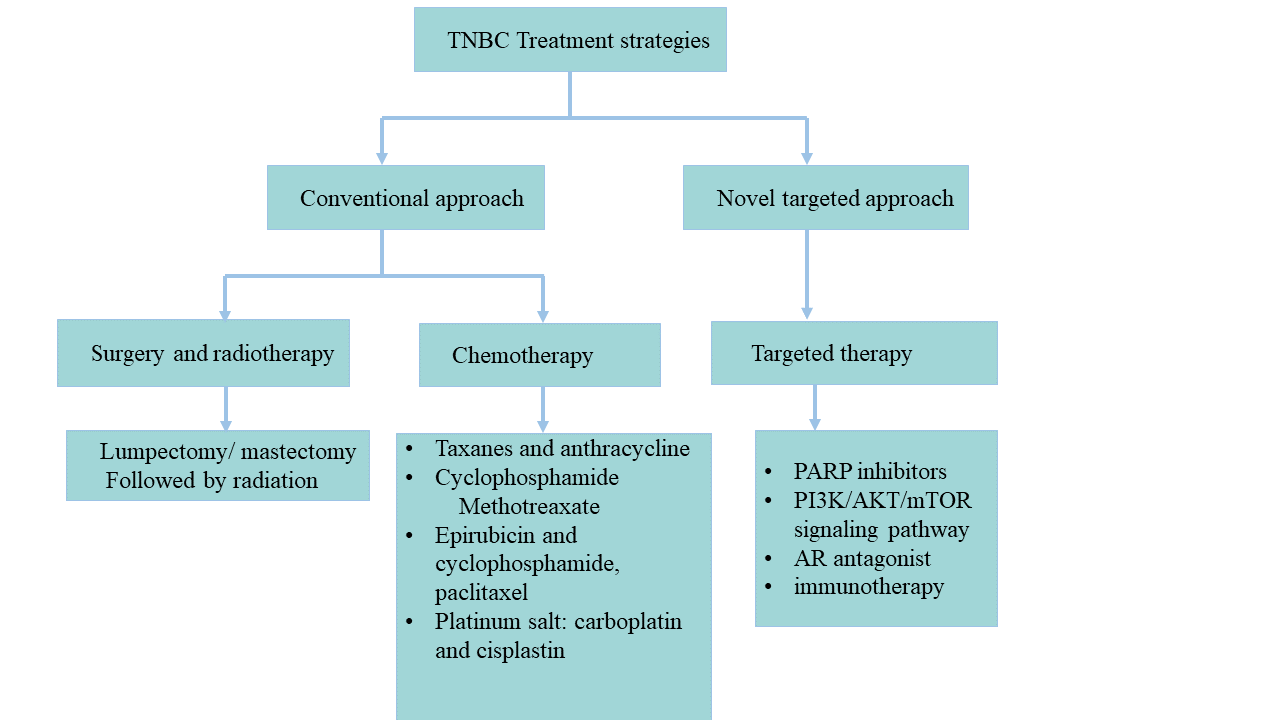
Figure 3. An Overview of conventional and new targeted approaches to treating triple-negative breast cancer
CONCLUSION
Triple-negative breast cancer (TNBC) remains one of the most challenging subtypes of breast cancer due to its molecular heterogeneity, aggressive clinical behavior, and lack of targeted therapies. The use of D-α-tocopheryl polyethylene glycol 1000 succinate (TPGS), a water-soluble derivative of vitamin E, presents a promising strategy to overcome these limitations. TPGS exhibits multifunctional therapeutic potential—not only as a nanocarrier that enhances drug solubility and cellular uptake but also as an active anticancer agent capable of inducing apoptosis, generating reactive oxygen species (ROS), and inhibiting P-glycoprotein-mediated drug efflux. These properties allow TPGS to synergize with conventional chemotherapeutic agents, improve their efficacy, and combat multidrug resistance in TNBC. Additionally, TPGS-based nanoformulations can be engineered to target specific signaling pathways commonly dysregulated in TNBC, such as PI3K/AKT/mTOR, Notch, Hedgehog, and Wnt/β-catenin. Collectively, these findings support the potential of TPGS as a versatile platform for the development of innovative therapeutic strategies against TNBC, offering a new avenue for effective and personalized cancer treatment.
REFERENCES

Indrajeet Yadav, Priyanka Sonker, TPGS Nano Formulation in Triple Negative Breast Cancer, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 6, 2464-2477. https://doi.org/10.5281/zenodo.15650192
 10.5281/zenodo.15650192
10.5281/zenodo.15650192
