Dept. of Pharmacology, Appasaheb Birnale College of Pharmacy, Sangli, Sangli, Maharashtra-416416.
The hormonal regulation of migraine pathology in women is influenced by fluctuations in key hormones such as estrogen, progesterone, oxytocin and prolactin, particularly during critical periods like pregnancy, menstruation, and menopause. This review examines the specific roles these hormones play in the modulation of migraine susceptibility and severity. Estrogen, with its significant fluctuations during the menstrual cycle, is known to be a major trigger for migraines, particularly in the premenstrual phase. Progesterone, which balances estrogen's effects, may have a protective role, especially during pregnancy, where the reduced frequency of migraines is observed. Oxytocin, often referred to as the "bonding hormone," may influence migraine patterns during pregnancy and in the postpartum period, though its direct involvement remains understudied. Prolactin, primarily associated with lactation, also shows potential links to migraine in the context of pregnancy and postpartum, possibly by affecting neurotransmitter systems. This review explores how the interplay of these hormones during pregnancy, menstruation, and menopause can provide insight into the pathophysiology of migraines and offers potential avenues for targeted therapeutic interventions for women suffering from hormone-related migraines.
Migraine is a prevalent neurovascular disorder characterized by recurrent moderate-to-severe headache attacks, with an estimated global prevalence of 15%(1) . Migraine is defined as severe throbbing plus unilateral headache related to nausea, photophobia, phonophobia and vomiting(2) The common forms of migraine - Migraine with/or without Aura (MA and MO, respectively), are diagnosed by the presence of recurrent headache that lasts 4-72 hours and is generally accompanied by nausea, photophobia, phonophobia, aggravation by physical activity and possible neurological symptoms, as outlined by the International Classification of Headache Disorders 2nd Edition (IHS 2004).(3)More than “just a headache” it is a disorder of central nervous system dysregulation that is three times more prevalent among women than men, with a cumulative lifetime prevalence among women of 43% (4) Many factors have been implicated in the pathogenesis of migraine headache, including activation of the trigeminovascular system, dysfunction of: cerebral blood vessels, circulating vasoactive substances, mitochondrial energy metabolism, brain oxygenation and metabolism, platelet disorder, alterations in serotonin levels, low levels of brain tissue magnesium, altered transport of ions across the cell membrane, and inheritance and dysfunction of the brainstem periaqueductal gray matter. The headache phase of migraine is associated with cerebral vasodilation and inflammation, presumably mediated by the release of vasoactive substances and neuropeptides including CGRP (calcitonin gene related peptide). Increased serum CGRP levels have been detected during migraine and cluster headache Migraine is a genetically influenced complex disorder characterized by episodes of moderate-to-severe headache, most often unilateral and generally associated with nausea and light and sound sensitivity. Migraine is predominantly a disorder of women that has long been linked with sex hormones. It poses a significant public health burden, particularly during the reproductive years.(5) Women’s reproductive cycle regulated by the hypothalamic hypophyseal-ovarian axis through the release of estrogen and progesterone. Variations in the levels of these hormones control and regulate the menstrual cycle, pregnancy, puerperium, and menopause.(6)
Hormonal Interactions in Migraine
The four hormones above do not act in isolation; they intersect in migraine pathway
Hormones Involved in Migraine Development
Estrogen
Estrogens are lipophilic hormones derived from cholesterol and synthetized primarily in the granulosa cells of ovaries. They also derive from the aromatization of androgens in peripheral tissues and within the brain.(7) The most relevant endogenous estrogen is E2, the classical female sex steroid hormone. Estrogens reach the central nervous system (CNS) passively diffusing through the blood–brain barrier, but they are also locally synthetized from cholesterol or converted from aromatizable precursors by the brain enzyme aromatase, thus acting as neurosteroids(8). The secretion of estrogens is regulated by bursts of gonadotropin-releasing hormone (GnRH) from the hypothalamus that in turn induce the release of FSH and LH from the pituitary.(9) Estrogens achieve physiological effects through the activation of various estrogen receptors (ERs), including three known forms: estrogen receptor-(ERα ), estrogen receptor-(ERβ), and the more recently identified G protein coupled estrogen receptor-1 (GPER/GPR30) (10). However, activation of different ERs presents different outcomes in pain modulation. For example, a study showed that ERβ agonists were effective in alleviating pain induced by chemotherapy, while the nonselective agonist 17β-estradiol and the ERα-selective agonist PPT had no effect in the same model [11). ERα predominates in the ventromedial hypothalamus but is absent in the cortex and cerebellum, whereas ERb is present in the cortex and cerebellum in addition to other regions, such as the preoptic area, amygdala, thalamus and some nuclei in the brain stem(8,12) Estrogen plays a significant role in modulating pain pathways, influencing pain perception and potentially contributing to pain conditions like migraine and fibromyalgia, with its effects mediated by estrogen receptors and other neural pathways. Because they are activated by ligands in either genetic or non-genetic ways, ERα and ERβ interact with the nociceptive system to modulate pain via different cellular signaling pathways(11). Low17-estradiol levels correlated with a spike in migraine attacks, and maximum percentage of the women experienced migraine attacks that were debilitating(12) Numerous areas involved in nociception in the nervous system contain ERα and/or β: DRG, dorsal horn of the spinal cord, periaqueductal gray, parabrachial nuclei, raphe nuclei,hypothalamus, limbic system, and several cortical areas.(13) Estrogen modulates pain perception through both genomic (nuclear estrogen receptors) and non-genomic (membrane-bound receptors) pathways, affecting neuronal activity and pain signaling in the central and peripheral nervous systems.(14).
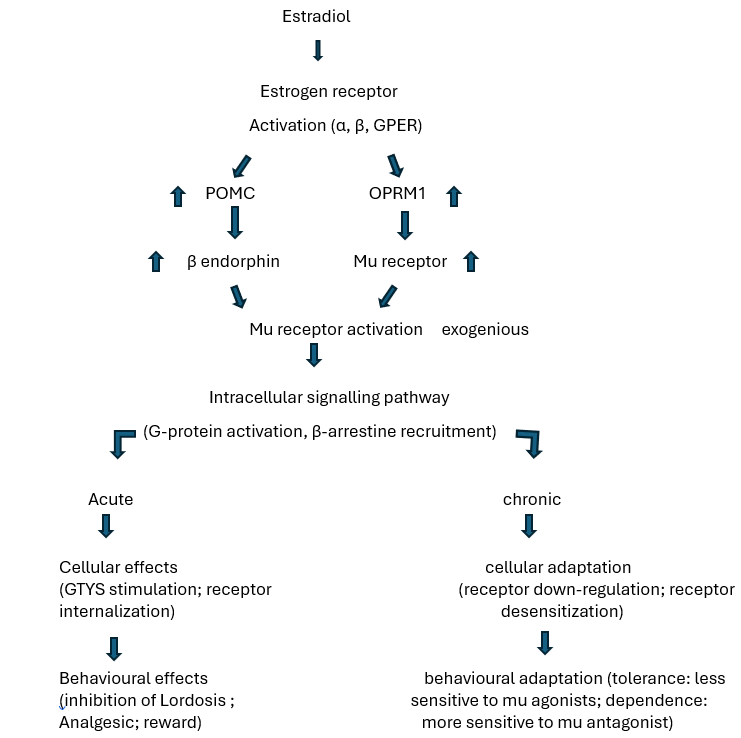
The effects of estradiol on mu opioid receptor endpoints under acute and chronic conditions. Estradiol binds and activates estrogen receptor subtypes (ER α , ERβ, and GPER). Activation of some or all of these receptors increases expression of the opioid receptor mu 1 (OPRM1) gene, which subsequently increases mu opioid receptor protein levels, and the proopiomelanocortin (POMC) gene, which subsequently increases β-endorphin peptide concentrations and mu receptor activation. Estradiol-induced activation of mu receptors via β-endorphin activates intracellular signalling pathways and leads to mu opioid receptor internalization and mu-mediated behavioural effects. Chronic estradiol treatment leads to compensatory down-regulation and desensitization of mu opioid receptors and changes in sensitivity to exogenous mu receptor agonists and antagonists. Estradiol functions as a high affinity agonist at the nuclear estrogen receptors alpha (ERα) and beta (ERβ), as well as the membrane-bound G-protein coupled estrogen receptor (GPER). These receptors are widely distributed across tissue types, and significant concentrations of these receptors are found in brain. Additionally, there is evidence that estradiol influences neurotransmission within the hippocampus via endogenous mu opioids(15). Estrogen also has an effect on the glutaminergic system which in turn has an impact on neural excitability within the trigeminal nucleus caudalis.
Neurotransmitter Interactions: Estrogen profoundly modulates key neurotransmitter systems:
Serotonin (5-HT):
E2 upregulates tryptophan hydroxylase and 5-HT receptor genes, and downregulates MAO-A/B and the serotonin transporter. The net effect is enhanced serotonergic tone (analgesic), which may protect against migraine. Notably, estrogen withdrawal reduces 5-HT availability, potentially triggering headache.
Glutamate:
Estrogen increases glutamatergic excitability (promoting new dendritic spines and glutamate receptor expression) High E2 states (e.g. pregnancy) thus raise cortical excitability and migraine aura risk.
GABA:
E2 enhances GABAergic inhibition (increasing GAD, GABA release and GABA_A receptor density), opposing nociception. Progesterone and its metabolite allopregnanolone similarly potentiate GABA_A function, producing an antinociceptive effect.
Opioids:
Estrogen upregulates endogenous opioids (e.g. enkephalins) and opioid receptor affinity, bolstering analgesia. Women with menstrual migraine may lose this opioid tone premenstrually.
CGRP:
Estrogen normally restrains CGRP. ERα activation suppresses pro-nociceptive mediators. Conversely, estrogen withdrawal (peri-menstrual drop) leads to a rebound increase in CGRP release. Immunohistochemical studies show that baseline CGRP release is higher in the proestrus (high-E2) rat state than in estrus, implying estrogen sets CGRP tone. Migraine attacks have been linked to low serotonin plasma levels [16]. Estrogens may increase the serotonergic tone and influence serotonin synthesis, reuptake and degradation [17]. This association might further contribute to the frequency of headache attacks when estrogen levels fall and consecutively serotonin plasma levels decrease as well. The calcitonin gene-related peptide (CGRP) is a sensory neuropeptide with a role in peripheral and central pain mechanisms including those leading to migraine. ERs are expressed in the same brain areas, including the TG, as CGRP receptors, thus indicating that estrogens can be involved in the modulation of CGRP signaling in the pathophysiology of migraine. Estrogen levels naturally fluctuate in women throughout their lives, particularly during puberty, the menstrual cycle, pregnancy, and menopause.
Fluctuation of estrogen during puberty:
During puberty, estrogen fluctuations, particularly the dip before menstruation, are strongly linked to migraine onset and frequency in women, as estrogen plays a key role in migraine pathophysiology. Migraine prevalence increases significantly in girls after puberty, coinciding with the onset of menstruation and the associated hormonal changes. (18) Estrogen levels fluctuate throughout the menstrual cycle, and a drop in estrogen, especially before menstruation, is a well-established trigger for migraine attacks. (19)
Fluctuation of estrogen during menstrual cycle Menstrual-related migraines happen in more than 50 percent of women who have migraine headaches. Menstrual migraine attacks usually last longer than other migraine attacks. (20) Migraines during menstruation, also known as menstrual migraines, are common and often linked to fluctuations in estrogen level. Estrogen levels rise during the mid-follicular phase and then drop precipitously after ovulation. This is followed by a secondary rise in estrogen levels during the mid-luteal phase with a decrease at the end of the menstrual cycle.(21) As many as 60% of female migraineurs report attacks during perimenstrual periods, Premenstrual headache occurs earlier in the cycle, typically 2 to 7 days before the onset of menses and may be part of premenstrual syndrome (PMS) The drop in estrogen during the luteal phase is believed to trigger migraine and this decline may impact blood vessels making them more permeable to pro-inflammatory mediators such as prostaglandins. Prostaglandin levels are elevated threefold in the luteal phase with a further increase during menstruation, and therefore thought to play a role in MRM(22) A drop in estrogen may cause an increased sensitivity to prostaglandins and a release of neuropeptides such as CGRP, substance P and neurokinins which could result in neurogenic inflammation [23]. This physiological response provokes alterations in the microvasculature of the dura mater, changes in calcium and magnesium concentrations, and an imbalance in serotonin and dopamine concentrations. Functional polymorphisms in estrogen metabolism genes COMT, CYP1A1, or CYP19A1 have not been found to be associated with MM. (24) However, several other hormone-related genes have been identified to be associated with predis position to migraine and, in particular, MM. These include ESR1, PRPROGINS insert, and possibly, ESR2 and FSHR, as well as genetic variants in the SYNE1 and TNF genes.
Fluctuation of estrogen during pregnancy:
During pregnancy, estrogen levels rise dramatically, peaking in the third trimester, and then plummet after birth. plasma levels of 17β-estradiol, estriol and progesterone increase to a level 10 to 100 times higher than that in the nonpregnant state. The majority of women suffering from migraine without aura and menstrual migraine experience improvement during pregnancy. After childbirth, estrogen levels drop rapidly to pre-pregnancy levels and remain suppressed during the postpartum period.(25)
Progesterone
Progesterone synthesis, a process called steroidogenesis, involves the conversion of cholesterol to pregnenolone, which is then converted to progesterone, primarily in the corpus luteum and placenta, and also in the brain. (26) Progesterone has been reported to be associated with anti-infammatory, anti-vasoconstriction, and neuroprotective functions.(27) Progesterone influences migraine through multiple mechanisms, particularly by modulating pain perception, neuroinflammation, and vascular stability.
1. Modulating Nociception : Progesterone, along with allopregnanolone (a progesterone metabolite), appears to dampen pain signals in the trigeminovascular system, a key pathway involved in migraine pain.(28)
2. Reducing Neurogenic Inflammation: Progesterone and allopregnanolone can reduce neurogenic inflammation, a process that contributes to migraine attacks, through interactions between neurons and glial cells. PR-B is particularly involved in neuroprotection by regulating gene expression related to cell survival, anti-inflammatory pathways, and neurotrophic factors. These receptors modulate inflammation by suppressing pro-inflammatory cytokines like IL-1, TNF-1, IL-6, and inflammatory signaling pathways like NF-KB.
3. Stabilizing the Blood-Brain Barrier: Progesterone helps stabilize the blood-brain barrier, which is crucial for protecting the brain from damage and maintaining its proper function. PGRMC1 involved in maintaining the blood-brain barrier (BBB) and reducing oxidative stress and apoptosis in neurons
4. Anti-inflammatory and Anti-apoptotic Effects: Progesterone has anti-inflammatory and anti-apoptotic (cell death) properties, which can help protect neurons and glial cells from damage. found behind the observed neuroprotection include reduced edema, reduced BBB permeability, and decreased expression of molecules associated with inflammation including IL-1β, TNF-α, IL-6, MCP-1, ICAM-1, NF-κB, and TLR4. PR-B is particularly involved in neuroprotection by regulating gene expression related to cell survival, anti-inflammatory pathways, and neurotrophic factors. PR-A and PR-B receptors modulate inflammation by suppressing pro-inflammatory cytokines like IL-1β, TNF-α, IL-6, and inflammatory signalling pathways like NF-κB. During pregnancy, progesterone plays a vital role in maintaining a healthy pregnancy by preparing the uterine lining for implantation, preventing contractions that could lead to preterm labor, and supporting the development of the foetus and breasts for breastfeeding. progesterone downregulate pro-inflammatory cytokines and/or to upregulate anti-inflammatory cytokines (29). Several immunological effects of progesterone are mediated by an immunomodulatory molecule secreted by pregnancy lymphocytes so-called progesterone-induced blocking factor (PIBF), a protein with inhibitory effects on cell-mediated immune reactions.(30) women with migraine generally experience decreased migraine symptoms by the third trimester. How ever, migraine can also start during pregnancy, and can worsen for women in the first trimester.(31) progesterone plays a role in preparing the uterus for pregnancy, and its fluctuations, particularly the drop before menstruation, may be linked to migraine attacks. Migraines linked to menstruation, often called "menstrual migraines," tend to occur during the late luteal phase, when estrogen and progesterone levels drop just before menstruation. The luteal phase, which follows ovulation, is characterized by high levels of progesterone and estrogen. However, these levels decline significantly just before menstruation, which is thought to be a trigger for menstrual migraines.
Oxytocin
Oxytocin(OT), a neuropeptide hormone, is primarily synthesized in the magnocellular neurons of the paraventricular (PVN) and supraoptic (SON) nuclei of the hypothalamus, and then transported to the posterior pituitary for release.(32) OT is also synthesized in peripheral tissues, e.g., uterus, placenta, amnion, corpus luteum, testis, and heart. The OT receptor is a typical class I G protein-coupled receptor that is primarily coupled via G(q) proteins to phospholipase C-beta.(33). OT, binding to trigeminal OTR, decreases the excitability of these trigeminal neurons so could contribute to decreased probability of a migraine attack. While there is only one main type of oxytocin receptor (OXTR), its function and signaling can vary depending on the cell type, tissue localization, and interacting proteins.
Types of oxytocin receptor signalling pathways
The β-arrestin signaling pathway plays a crucial role in migraine pathophysiology, particularly in how pain is processed, how receptors desensitize, and how inflammation is regulated in the brain.
β-arrestin are scaffold proteins that regulate GPCR activity in two major ways:
1. Desensitization & Internalization of GPCRs
? After repeated activation, GPCRs (such as serotonin or oxytocin receptors) become desensitized (less responsive). Desensitization of G-protein-coupled receptors (GPCRs) through β-arrestin recruitment plays a dual role in migraine management. It can be beneficial or problematic, depending on the receptor type and drug being used. Certain receptors contribute to migraine pain when they remain overactivated, excessive β-arrestin recruitment can reduce drug effectiveness, leading to tolerance and rebound headaches. Desensitization via β-arrestin can help reduce migraine severity by limiting excessive signaling in these pathways.e.g Chronic triptan use leads to 5-HT1B/1D receptor desensitization, making the drug less effective over time. While controlled desensitization is beneficial. When CGRP receptors (CGRP-R) are desensitized, they become less responsive to CGRP, reducing migraine frequency and severity. CGRP inhibitors (Gepants like rimegepant, ubrogepant) promote desensitization of CGRP-R without excessive β-arrestin recruitment, avoiding tolerance. β-arrestins help internalize these receptors, temporarily stopping their activity. Internalization refers to the process where a G-protein-coupled receptor (GPCR) is removed from the cell surface and transported inside the cell, preventing it from being activated by ligands. β-arrestin recruits clathrin and adaptor proteins, forming a clathrin-coated vesicle. The receptor is pulled inside the cell in an endosome, removing it from the cell membrane so it cannot be activated.
2. G-Protein-Independent Signalling
Studies on menstrual migraine have shown that estrogen regulates OT release and the expression of the OTR (34)During menstruation there is a reduction of estrogen and OT, as well as a reduction of magnesium and cholesterol, which positively modulate the affnity of OT for OTRs. Consequences are a decrease of OT levels, reduced affnity of OT for its receptor and a decreased expression of the trigeminal OTR. All of these variations seem to determine activation of meningeal trigeminal nociceptors and increase the risk of menstrually related migraine attacks, suggesting trigeminal OTR as a therapeutic target for menstrually related migraine.(35) OT is increased during pregnancy, coinciding with a general decrease of headaches and migraine during pregnancy More specifically, this reduction in headache occurs in patients with migraine without aura, in which case there is an increase in the frst trimester and a decrease in the following two trimesters, but new onset aura may appear at that time.(36) OT activates the OTR resulting in intracellular mobilisation of Ca2+, thereby inhibiting nociception by different actions: GABAergic signalling, inhibition of transient potassium current, desensitisation of spinal TRPV1 channels, and disruption of the NMDA-evoked coordinated neuronal network activity [37). Assessing the anatomical side, the anti-migraine action follows two different pathways: projection from the hypothalamic paraventricular nucleus to the nucleus caudalis and trigeminal nociceptive pathways via activation of the OTRs.
Prolactin
Prolactin (PRL) is a polypeptide hormone expressed at lactotrophic cells in the anterior pituitary gland. PRL is a member of the prolactin/growth hormone/placental lactogen family 1 with 199 amino acids arranged in loops and alpha helixes. (38) Prolactin plays regulatory and modulatory roles in the endocrine, immune and nervous systems and has been associated with the etiology of pain and headaches . Prolactin secretion is influenced by many factors, including physiologic factors (e.g., pregnancy, sleep and stress), pathologic factors (e.g., pituitary adenoma, hypothyroidism and renal failure), and some drugs (e.g., antipsychotics, antidepressants, prokinetics, morphine and antihypertensive agents). Prolactin levels follow a circadian rhythm ,adjusted by the hypothalamus, which is also believed to be involved in the onset of migraines . Premonitory symptoms (e.g.,food craving and mood swing) have been linked to hypothalamic dysfunction , and perturbations in the hypothalamic–pituitary–gonadal (HPG) axis have been associated with menstrual migraine.(39) PRL contributes to the modulation of sensory neurons in the trigeminovascular system, the physiological substrate of migraine pathogenesis. Recent data demonstrated that PRL administration induced migraine- like behaviour in rodents, indicating involvement of PRL in cellular cascades leading to migraine attacks. These observations and the findings that PRL signalling is involved in sex- related differences in pain perception might explain the sexual dimorphic etiology of migraine.(40) It should be noted that interoceptive physiological processes that disrupt homeostasis, such as sleep disruption, missing meals, fluid imbalance and others that are often identified as‘migraine triggers’,could also engage hypothalamic kappa opioid receptor (KOR) circuits to promote nociceptor sensitization through this neuroendocrine mechanism. Similarly, psychological stress that arises from daily life and is most commonly perceived as a migraine trigger probably influences hypothalamic activity.(41) The mechanisms by which increased levels of circulating PRL and dysregulation of PRLR isoforms could result in increased likelihood of migraine in females remain to be elucidated. However, it is known that PRL promotes sensitization of responses to activation of TRPV1, TRPM8 and TRPA1-positive sensory neurons selectively in female DRG and TG neurons. Our studies provoked allodynia with a subthreshold exposure to UMB, a TRPA1 agonist, suggesting the possibility of modulation of TRPA1 expressing fibres. Furthermore, activation of trigeminal TRP channels increases the release of CGRP, a peptide known to promote migraine in humans.(42) The long-form PRLR activates JAK2–STAT signaling (driving transcriptional changes), while short-form receptors preferentially engage MAPK and PI3K pathwaysPRLR signaling can also cross-talk with MAPK/ERK pathways, affecting cell survival and functionPRL’s downstream genes include those involved in immune modulation and neuroendocrine function.
Neurotransmitter and Ion Channel Interactions: In the trigeminovascular system, PRL is pronociceptive. It sensitizes primary afferents by modulating ion channels and neuropeptide release. Key mechanisms (from recent studies) include:
Table 1: hormones, receptors and localization with signalling pathway.
|
Hormone |
Receptor(s) & Localization |
Signalling Pathways |
|
Estrogen |
ERα, ERβ (nuclear; e.g. trigeminal ganglion nuclei) GPER (membrane) |
Genomic: ERα/β → transcription of genes (ion channels, cytokines) Non-genomic: GPER (Gq → ↑Ca²?, Gi → ↓cAMP) |
|
Progesterone |
PR-A, PR-B (nuclear); membrane PR (mPR) |
Genomic: PR → transcription of anti-inflammatory/neuroprotective genes. Metabolites (allopregnanolone) → modulate GABA_A receptors (non-genomic). |
|
Oxytocin |
OTR (GPCR) – widely in CNS/PNS (brainstem, spinal trigeminal tract, trigeminal ganglia) |
GPCR (Gq → ↑Ca²?; Gi → ↓cAMP). Triggers intracellular Ca²? signaling that recruits GABAergic and peptidergic inhibition. |
|
Prolactin |
PRLR (class I cytokine receptor; long and short isoforms) |
Long PRLR: JAK2 → STAT transcription factors. Short PRLR: activates MAPK/ERK and PI3K cascades |
CONCLUSION
This review highlights how cyclical sex steroids modulate migraine in women. Migraine is roughly three times more prevalent in women and closely tied to reproductive events: premenstrual estrogen withdrawal often precipitates attacks, whereas the sustained high estrogen of pregnancy or postmenopausal hormone decline tend to ameliorate them Estradiol influences key neuromediators – for example, it shapes serotonergic signalling and CGRP release. Progesterone may counterbalance these effects: progestogen-only contraceptive regimens often reduce menstrual migraine frequency. Pituitary prolactin and hypothalamic oxytocin add another layer of female-specific modulation. PRL is pronociceptive – sensitizing trigeminal TRPV1 (a pain-sensing TRP channel)– whereas OT is largely analgesic Notably, some trigeminal neurons co-express CGRP and oxytocin receptors, suggesting a neuroendocrine push–pull between migraine-promoting and suppressing signals. These converging findings underscore that reproductive endocrine fluctuations uniquely shape women’s migraine patterns and burden. Future studies should clarify oxytocin and prolactin mechanisms and leverage them for targeted, sex-specific therapies (e.g. oxytocin analogues or prolactin antagonists).
ACKNOWLEDGEMENT:
I extend my sincere gratitude to my guide, Dr. Smt .N. S. Naikwade, for her expert guidance and invaluable contributions to this work. I am also deeply thankful to my teachers for their insightful feedback and constant encouragement. Additionally, I appreciate the unwavering support and constructive discussions from my friends, which greatly enriched this review. Their collective efforts were instrumental in the completion of this article.
REFERENCES




Minal Kumbhar*, Aarti Mole, Dr. N. S. Naikwade, Janhavi Mali, Hormonal Fluctuations and Their Impact on Migraine Susceptibility in Women, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 6, 4233-4245. https://doi.org/10.5281/zenodo.15736654
 10.5281/zenodo.15736654
10.5281/zenodo.15736654
