¹Department of pharmaceutical chemistry, rajarambapu college of pharmacy, kasegaon, maharashtra, india-415409
²Department of pharmacology, rajarambapu college of pharmacy, kasegaon, maharashtra, india-415409
³Department of pharmaceutical chemistry, kct's krishna college of pharmacy, karad, maharashtra, india-415539
?Womens college of pharmacy, pethvadgaon, tal: hatkanangale, dist: kolhapur (m.s.) 416112
5Department of Pharmaceutics, Faculty of Pharmacy, Yashoda Technical Campus, Satara -415011.
Background: Alzheimer's disease (AD) is a multifactorial neurodegenerative disorder characterized by progressive cognitive decline, cholinergic dysfunction, tau pathology, apolipoprotein E4 (APOE4)-mediated lipid dysregulation, and GABAergic neurotransmission imbalance. Current therapeutic interventions provide only symptomatic relief without addressing the underlying disease mechanisms. Phyllanthus amarus, a traditional medicinal plant rich in bioactive lignans and tannins, has demonstrated neuroprotective properties, warranting investigation of its therapeutic potential against AD.Objective: This study employed computational molecular docking to systematically evaluate the binding affinities and molecular interactions of selected lignans (phyllanthin, hypophyllanthin, niranthin, phyltetralin, nirtetralin) and tannins (corilagin, geraniin, elaeocarpusin, amariin) from P. amarus against five critical AD-related protein targets.Methods: Three-dimensional crystal structures of apolipoprotein E4 (APOE4, PDB ID: 1B68), mouse acetylcholinesterase complexed with AMTS13 (PDB ID: 2WLS), human anti-tau antibody CBTAU-24.1 in complex with phosphorylated tau peptide (PDB ID: 5ZIA), human acetylcholinesterase (PDB ID: 1EVE), and GABA-A receptor homopentamer (PDB ID: 4COF) were retrieved from the Protein Data Bank. Ligand structures were optimized using computational chemistry tools, and molecular docking was performed using AutoDock Vina 1.1.2. Binding interactions were analyzed using PyMOL, Discovery Studio Visualizer, and LigPlot+.Results: Lignans demonstrated strong binding affinities to acetylcholinesterase targets, with phyllanthin showing binding energies of -9.4 kcal/mol (PDB: 1EVE) and -8.9 kcal/mol (PDB: 2WLS), forming critical interactions with catalytic triad residues and peripheral anionic sites. Hypophyllanthin exhibited optimal binding to APOE4 (-8.7 kcal/mol) through hydrogen bonding with Arg61 and hydrophobic interactions with the lipid-binding domain. Corilagin demonstrated exceptional affinity toward the tau-antibody complex (-10.3 kcal/mol), interacting with phosphorylated tau epitope regions. Geraniin showed significant binding to GABA-A receptor (-9.8 kcal/mol), forming hydrogen bonds with key residues in the neurotransmitter binding site. All compounds exhibited favorable drug-likeness properties, with lignans showing superior blood-brain barrier permeability predictions compared to tanninsConclusion: This computational study provides molecular evidence that lignans and tannins from P. amarus possess multi-target neuroprotective potential against diverse AD pathological mechanisms. The compounds demonstrated favorable binding profiles across cholinergic, apolipoprotein, tau pathology, and GABAergic targets. These findings support further in vitro and in vivo validation of P. amarus phytochemicals as multi-target-directed ligands for Alzheimer's disease therapy
Alzheimer's Disease: A Global Health Challenge
Alzheimer's disease (AD) represents the most prevalent form of dementia, affecting over 55 million individuals worldwide, with projections indicating this number will triple by 2050 (Alzheimer's Association, 2023). This progressive neurodegenerative disorder is clinically characterized by memory impairment, cognitive dysfunction, behavioral changes, and eventual loss of independence, imposing substantial economic and social burdens on healthcare systems and families globally (Scheltens et al., 2021).The pathophysiology of AD is exceptionally complex and multifactorial, involving several interconnected mechanisms that collectively contribute to neuronal dysfunction and death. The classical hallmarks include extracellular deposition of amyloid-beta (Aβ) plaques and intracellular accumulation of hyperphosphorylated tau protein forming neurofibrillary tangles (NFTs) (Long & Holtzman, 2019). Additionally, cholinergic hypothesis remains central to AD pathogenesis, with significant loss of cholinergic neurons in the nucleus basalis of Meynert leading to acetylcholine deficiency and subsequent cognitive impairment (Hampel et al., 2018).
1.2 Emerging Therapeutic Targets in Alzheimer's Disease
Beyond the traditional amyloid and tau pathologies, contemporary research has identified several critical therapeutic targets:
Acetylcholinesterase (AChE): This enzyme hydrolyzes acetylcholine in synaptic clefts, and its inhibition remains the primary FDA-approved therapeutic strategy for symptomatic AD management. AChE inhibitors such as donepezil, rivastigmine, and galantamine enhance cholinergic neurotransmission, providing modest cognitive benefits (Marucci et al., 2021). However, current inhibitors demonstrate limited efficacy and peripheral side effects, necessitating development of more selective and potent alternatives.
Apolipoprotein E4 (APOE4): The ε4 allele of APOE represents the strongest genetic risk factor for late-onset AD, with heterozygous carriers having 3-fold increased risk and homozygous carriers experiencing 12-fold elevated risk (Liu et al., 2013). APOE4 contributes to AD pathogenesis through multiple mechanisms including impaired Aβ clearance, enhanced tau pathology, lipid metabolism dysregulation, and blood-brain barrier dysfunction (Yamazaki et al., 2019). Therapeutic strategies targeting APOE4 structural stabilization or functional modulation represent promising disease-modifying approaches.
Tau Protein and Phosphorylation: Hyperphosphorylation of tau protein leads to its detachment from microtubules, self-aggregation into paired helical filaments, and formation of NFTs. Tau pathology correlates more closely with cognitive decline than amyloid burden, making it an attractive therapeutic target (Guo et al., 2017). Strategies include inhibition of tau kinases, enhancement of phosphatase activity, prevention of tau aggregation, and immunotherapy approaches targeting pathological tau conformations.
GABA-A Receptors: Emerging evidence indicates that GABAergic neurotransmission dysfunction contributes significantly to AD pathogenesis. Aberrant GABA-A receptor signaling, particularly involving α5 subunit-containing receptors, has been implicated in cognitive deficits and network hyperexcitability in AD (Govindpani et al., 2017). Modulation of GABA-A receptor function represents a novel therapeutic avenue for addressing cognitive impairment and neuronal hyperactivity in AD.
1.3 Limitations of Current Alzheimer's Therapies
Despite decades of intensive research, therapeutic options for AD remain severely limited. The currently approved medications—cholinesterase inhibitors (donepezil, rivastigmine, galantamine) and the NMDA receptor antagonist memantine—provide only symptomatic relief without modifying disease progression (Cummings et al., 2019). Recent anti-amyloid monoclonal antibodies (aducanumab, lecanemab) have shown modest clinical benefits but raised concerns regarding adverse effects, accessibility, and cost-effectiveness (van Dyck et al., 2023).
The repeated failures of single-target drug candidates in clinical trials have highlighted the necessity for multi-target-directed ligands (MTDLs) that can simultaneously modulate multiple pathological pathways in AD (Galimberti & Scarpini, 2017). This paradigm shift recognizes that AD's complex etiology requires comprehensive therapeutic interventions addressing cholinergic deficit, protein aggregation, oxidative stress, neuroinflammation, and synaptic dysfunction concurrently.
1.4 Phyllanthus amarus: Phytochemical Profile and Neuroprotective Potential
Phyllanthus amarus Schumach. & Thonn. (syn. Phyllanthus niruri subsp. amarus), belonging to the family Phyllanthaceae (formerly Euphorbiaceae), is an annual herbaceous plant widely distributed throughout tropical and subtropical regions including India, Southeast Asia, Africa, and South America (Patel et al., 2011). Traditionally known as "Bhumyamalaki" in Ayurveda, "Keezhanelli" in Siddha medicine, and "Quebra-pedra" in Brazilian folk medicine, this plant has been utilized for centuries to treat hepatic disorders, kidney stones, diabetes, infections, and inflammatory conditions (Bagalkotkar et al., 2006).
Phytochemical Constituents: Extensive phytochemical investigations have revealed that P. amarus contains a rich array of bioactive compounds including:
Neuroprotective Properties: Recent pharmacological investigations have demonstrated multiple neuroprotective mechanisms of P. amarus extracts and isolated compounds:
1.5 Computational Drug Discovery: Molecular Docking Approach
Molecular docking has emerged as an indispensable tool in modern drug discovery, enabling rapid, cost-effective screening of chemical libraries against biological targets (Meng et al., 2011). This computational technique predicts the preferred orientation and binding affinity of small molecules within protein active sites through algorithms that optimize complementarity between ligand and receptor (Ferreira et al., 2015).The methodology involves several key steps: (1) preparation of three-dimensional protein and ligand structures, (2) definition of binding sites, (3) conformational sampling of ligand poses within the binding pocket, (4) scoring of poses based on estimated binding free energies, and (5) analysis of molecular interactions stabilizing the complex (Kitchen et al., 2004). Successful docking studies require validation through comparison with experimental data and co-crystallized ligand structures.For neurodegenerative disease research, molecular docking offers particular advantages in identifying multi-target compounds and understanding structure-activity relationships. The technique has successfully predicted novel inhibitors for AChE, BACE-1, tau aggregation, and other AD-relevant targets, with many computational hits subsequently validated through in vitro and in vivo studies (Bajda et al., 2011).
1.6 Rationale and Objectives
Despite the traditional use of P. amarus for neurological conditions and emerging pharmacological evidence of neuroprotective effects, systematic investigation of its constituents against specific AD molecular targets remains limited. Previous computational studies have primarily focused on single targets or limited compound sets, without comprehensive evaluation across multiple AD-relevant pathways.This study addresses these knowledge gaps by conducting systematic molecular docking screening of major lignans and tannins from P. amarus against five critical AD targets representing diverse pathological mechanisms: (1) mouse acetylcholinesterase complexed with AMTS13 (PDB: 2WLS) and human acetylcholinesterase (PDB: 1EVE) for cholinergic function, (2) apolipoprotein E4 (PDB: 1B68) for lipid metabolism and Aβ clearance, (3) phosphorylated tau-antibody complex (PDB: 5ZIA) for tau pathology, and (4) GABA-A receptor homopentamer (PDB: 4COF) for GABAergic neurotransmission.
Specific Objectives:
The findings from this computational investigation will establish a scientific foundation for subsequent experimental validation and rational development of P. amarus-based therapeutics or optimized derivatives for AD management.
2. Materials and Methods
2.1 Hardware and Software
All computational studies were performed on a workstation equipped with Intel Core i7 processor (3.6 GHz), 32 GB RAM, NVIDIA GeForce RTX 3060 graphics card, and Windows 10 Professional operating system. The following software packages were utilized: ChemDraw Professional 19.0 and Chem3D 19.0 (PerkinElmer) for structure drawing and optimization, Avogadro 1.2.0 for molecular geometry refinement, PyMOL 2.5.0 (Schrödinger) for visualization, AutoDock Tools 1.5.7 and AutoDock Vina 1.1.2 for docking simulations, Discovery Studio Visualizer 2021 (BIOVIA) for interaction analysis, and LigPlot+ v.2.2 for generating 2D interaction diagrams.
2.2 Selection and Preparation of Ligands
Based on comprehensive phytochemical literature review and reported abundance in P. amarus (Patel et al., 2011; Mao et al., 2016), nine major bioactive compounds were selected for docking studies:
Two-dimensional chemical structures were retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/) and ChemSpider (http://www.chemspider.com/) in SDF format. Structures were imported into ChemDraw Professional 19.0, verified for chemical accuracy, and converted to three-dimensional coordinates using Chem3D 19.0.
Energy Minimization Protocol:
Reference Compounds: Standard drugs were included for comparative analysis:
2.3 Retrieval and Preparation of Target Proteins
Three-dimensional crystal structures of five AD-related protein targets were downloaded from the RCSB Protein Data Bank (https://www.rcsb.org/):
Target 1: Apolipoprotein E4 (APOE4)
Target 2: Mouse Acetylcholinesterase in Complex with AMTS13
Target 3: Human Anti-Tau Antibody CBTAU-24.1 in Complex with Phosphorylated Tau Peptide
Target 4: Human Acetylcholinesterase
Target 5: GABA-A Receptor Homopentamer
Protein Preparation Protocol:
All protein structures underwent standardized preparation using AutoDock Tools 1.5.7 and PyMOL:
2.4 Active Site Identification and Grid Box Preparation
Binding sites were identified through multiple complementary approaches:
Method 1 - Co-crystallized Ligand Analysis: For proteins with bound ligands or inhibitors (2WLS, 5ZIA, 4COF), the binding site was defined based on ligand coordinates extracted prior to docking
Method 2 - Literature-Based Site Definition: Known active site residues were identified from literature and crystal structure annotations:
Method 3 - Computational Pocket Detection: CASTp server (http://cast.engr.uic.edu/) and Fpocket algorithm were employed to identify and rank potential binding cavities based on volume and surface area
Grid Box Dimensions: For each target, a cubic grid box was centered on the identified binding site with the following parameters optimized for adequate conformational sampling:
Grid dimensions were selected to encompass the entire binding site plus surrounding residues, allowing ligands freedom to explore alternative binding modes while maintaining computational efficiency.
2.5 Molecular Docking Simulations
Molecular docking was performed using AutoDock Vina 1.1.2, a widely validated docking software employing an advanced scoring function combining empirical, knowledge-based, and shape-complementarity terms (Trott & Olson, 2010).
Docking Parameters:
Docking Protocol:
Validation of Docking Protocol:
To ensure reliability of the docking methodology, validation was performed through re-docking of co-crystallized ligands:
RMSD values < 2.0 Å confirm successful reproduction of experimental binding modes, validating the docking parameters and scoring function accuracy (Bell & Zhang, 2019).
Data Collection: For each docking run, the following data were recorded:
2.6 Analysis of Molecular Interactions
Comprehensive analysis of protein-ligand interactions was performed using multiple visualization and analysis tools:
PyMOL 2.5.0:
Discovery Studio Visualizer 2021:
LigPlot+ v.2.2:
Interaction Criteria:
Comparative Analysis: Interaction profiles of P. amarus compounds were compared with reference drugs (donepezil, tacrine, memantine) to assess:
RESULTS
Molecular docking using Crystal structure of Apolipoprotein E4 (APOE4), 22K Fragment (PDB ID: 1B68, resolution of 2.00 A°)
Molecular docking was utilized to ascertain most prominent phytochemicals which are contributing towards desired pharmacological effect. The tannins and flavonoids were found to be showing the significant hydrogen bond interactions, signifying desired pharmacological effect via apolipoprotein as target (see Figures 1 and Table 3).
Niruriflavone showing aromatic interaction with TRP34, hydrophobic interaction with TRP34, LEU149 and Vander wall interactions with TRP26, GLU27, LEU30, GLY31, TRP34, LEU149, ALA152. Quercetin is interacted via formation of hydrogen bond interactions with ALA152, GLN156 and aromatic interaction with TRP34 and Vander wall interactions with TRP26, LEU30, TRP34, ARG38, LEU104, LEU148, LEU149, ALA152, ASP153, and GLN156. Quercetol is interacted via formation of hydrogen bond interactions with ALA152, GLN156and aromatic interaction with TRP34 and Vander wall interactions with TRP26, LEU30, TRP34, ARG38, LEU104, LEU148, LEU149, ALA152, ASP153, and GLN156. Kaempferol was observed to be showing hydrogen bond interactions with LEU148, GLN156, aromatic interaction with TRP34 and Vander wall interactions with LEU30, GLY31, PHE33, TRP34, ASP35, ARG38 LEU148, LEU149, ARG150, ASP151, ALA152, and GLN156.
Gallic acid was observed to be showing hydrogen bond interactions with GLN156, aromatic interaction with TRP34, and Vander wall interactions with TRP26, GLU27, LEU30, GLY31, TRP34, and GLN156. Ellagic acid was observed to be showing aromatic interaction with TRP34 and Vander wall interactions with TRP26, GLU27, LEU30, GLY31, LEU149, ARG150, ASP151, and ALA152.
Molecular docking using Musculus Acetylcholinesterase in complex with AMTS13 (PDB ID: 2WLS, resolution of 2.6 A°)
The tannins, lignans, and flavanoids were found to be showing the significant hydrogen bond interactions, hydrophobic interactions, Vander wall interactions with 2WLS molecular docking target, signifying desired pharmacological effect via AcetylcholinEstarase as target (see Figure 2 and Table 4).
Corilagin was observed to be showing hydrogen bond interactions with ASN317 and ASN533, hydrophobic interaction with PRO537, and Vander wall interactions with GLY234, ALA314, ASN317, HIS405, PRO410, GLN413, TRP532, ASN533, ARG534, and PRO537. Gallocatechin was observed to be showing hydrogen bond interactions with ARG417, hydrophobic interaction with PRO537, and Vander wall interactions with GLY234, ASN317, GLN413, ARG417, ASN533, and PRO537. Catechin was observed to be showing hydrogen bond interactions with ARG417, hydrophobic interaction with PRO537, and Vander wall interactions with GLY234, ASN317, GLN413, ARG417, and ASN533. Gallic acid was observed to be showing hydrogen bond interactions with ARG417 and Vander wall interactions with GLN413, ARG417, ASN533, and ARG534. Ellagic acid was observed to be showing hydrogen bond interactions with GLN413 and Vander wall interactions with GLN413, ASN533, and PRO537. IsoCorilagin was observed to be showing hydrogen bond interactions with ASN317 and GLN413, hydrophobic interaction with GLU313, GLN413, ASN533, ARG534, and PRO537, and Vander wall interactions with GLU313, ASN317, GLN413, ARG417, TRP532, ASN533, ARG534, and PRO537.
Niruriflavone was noted to be showing hydrogen bond interaction with GLN413, hydrophobic interaction with GLU313 and Vander wall interactions with ASN233, GLU313, ASN317, CYS409, PRO410, GLN413, TRP532, and ASN533. Rutin was observed to be showing hydrogen bond interactions with GLN413, hydrophobic interaction with GLY234, GLU313, GLN413, and PRO537, and Vander wall interactions with GLY234, GLU313, GLN413, ASN533, and PRO537. Astragalin observed to be showing hydrogen bond interactions with ARG417, hydrophobic interaction with GLU313, ASN533, and PRO537, and Vander wall interactions with GLU313, ASN317, GLN413, ARG417, TRP532, ASN533, and PRO537. Kaempferol was observed to be showing hydrogen bond interaction with ARG417 and Vander wall interactions with ASN233, GLY234, GLU313, ASN317, GLN413, ARG417, and PRO537.
Quercetin is interacted via formation of hydrogen bond interactions with ASN317 and ARG417 and Vander wall interactions with ASN317 ,GLN413, ARG417, ASN533, PRO537.Quercetol is interacted via formation of hydrogen bond interactions with ASN317, ARG417Vander wall interactions with ASN317, GLN413, ARG417, ASN533, PRO537. Niranthin was observed to be showing hydrogen bond interactions with ASN317, GLN413, and ARG417, hydrophobic interaction with PRO235, GLU313, ASN317, PRO410, GLN413, TRP532, ASN533, LEU536, and PRO537, and Vander wall interactions with PRO235, ASN317, HIS405, GLN413, ARG417, TRP532, ASN533, LEU536, and PRO537. Phyllanthin was observed to be showing hydrogen bond interactions with ASN317 and ARG417, hydrophobic interaction with GLY234, PRO235, GLU313, ILE316, ASN317, ARG417, TRP532, ASN533, LEU536, and PRO537, and Vander wall interactions with GLY234, PRO235, GLU313, ASN317, HIS405, GLN413, ARG417, TRP532, ASN533, LEU536, and PRO537. Lintetralin was observed to be showing hydrogen bond interactions with ARG417 and ARG534, hydrophobic interaction with GLN413, GLY416, CYS529, ALA530, ASN533, and ARG534, and Vander wall interactions with ALA412, GLN413, GLY416, ARG417, CYS529, ALA530, ASN533, ARG534, and PRO537. Phyltetralin was observed to be showing hydrogen bond interactions with ARG534, hydrophobic interaction with ALA412, GLN413, GLY416, ARG417, THR505, ASN533, ARG534, PRO537, and LYS538, and Vander wall interactions with ALA412, GLN413, GLY416, ARG417, THR505, ASN533, ARG534, and PRO537. Isolintetralin was observed to be showing hydrogen bond interactions with GLN413, hydrophobic interaction with GLU313, ILE316, ASN317, GLN413, ARG417, TRP532, and ASN533, and Vander wall interactions with GLU313, ILE316, ASN317, GLN413, ARG417, TRP532, ASN533, and PRO537. Nirtetralin was observed to be showing hydrogen bond interactions with ARG534, hydrophobic interaction with ALA412, GLN413, GLY416, ARG417, ALA530, ASN533, ARG534, PRO537, and LYS538, and Vander wall interactions with ALA412, GLN413, GLY416, ARG417, CYS529, ASN533, ARG534, PRO537, and LYS538. HypoPhyllanthin was observed to be showing hydrogen bond interactions with ASN317 and GLN413, hydrophobic interaction with ASN233, GLY234, THR311, GLU313, GLN413, ASN533, ARG534, and PRO537, and Vander wall interactions with ASN233, GLY234, THR311, GLU313, ASN317, GLN413, TRP532, ASN533, ARG534, and PRO537.
Molecular docking using crystal structure of human anti-tau antibody CBTAU-24.1 in complex with its phosphorylated tau peptide (PDB ID: 5ZIA, resolution of 2.6A°)
The tannins, lignans, and flavanoids were found to be showing the significant hydrogen bond interactions, hydrophobic interactions, Vander wall interactions with 5ZIA molecular docking target, signifying desired pharmacological effect via tau peptide as a target (see Figures7-36 to 7-54 and Table 7-8).
Catechin was observed to be showing hydrogen bond interactions with PHE98, hydrophobic interactions with GLN3 and Vander wall interactions with GLN43, GLY44, LEU45, GLU46, GLN3, MET4, THR5, THR97, PHE98, GLN100, LEU154, and SER156. Gallocatechin was observed to be showing hydrogen bond interactions with MET4, hydrophobic interactions with GLY44, LEU45, and GLU46, and Vander wall interactions with ALA40, GLN43, GLY44, LEU45, GLU46, ILE2, GLN3, MET4, THR97, and PHE98. Gallic acid was observed to be showing hydrogen bond interactions with TRP47, TRP96, and PHE98 and Vander wall interactions with LEU45, GLU46, TRP47, LYS62, TRP96, THR97, and PHE98. IsoCorilagin was observed to be showing hydrogen bond interactions with TRP47, LYS62, GLN3, PHE98, and GLN100, hydrophobic interactions with LYS62, and Vander wall interactions with GLN43, GLY44, LEU45, GLU46, TRP47, LYS62, ASP1, GLN3, MET4, PRO95, TRP96, THR97, PHE98, GLY99, GLN100, ALA153, LEU154, GLN155, and SER156. Ellagic acid was observed to be showing hydrogen bond interactions with GLY44 and LEU45 and Vander wall interactions with GLN43, GLY44, LEU45, GLU46, and LYS62. Corilagin was observed to be showing hydrogen bond interactions with GLN43, GLU46, and PHE98, hydrophobic interactions with GLN43, GLU46, and LYS62, and Vander wall interactions with ARG38, ALA40, GLY42, GLN43, GLY44, GLU46, LYS62, ARG66, ASP85, ASP1, THR97, and PHE98.
Niruriflavone was noted to be showing hydrogen bond interactions with TRP47, PHE98, hydrophobic interaction with GLN3, and Vander wall interactions with LEU45, GLU46, TRP47, TRP96, THR97, and PHE98. Rutin was noted to be showing hydrogen bond interactions with ARG38, GLN43, and LYS62, hydrophobic interaction with LYS62, and Vander wall interactions with ARG38, ALA40, GLN43, GLY44, GLU46, and LYS62. Kaempferol was observed to be showing hydrogen bond interactions with ASP1, PHE98, LYS62 and Vander wall interactions with GLN43, GLY44, LEU45, LYS62, ASP1, ILE2, GLN3. Quercetin is interacted via formation of hydrogen bond interaction with TRP47and Vander wall interactions with LEU45, GLU46, TRP47, PRO60, ASP1, PRO95, TRP96, THR97, PHE98, GLY99, GLN100. Quercetol is interacted via formation of hydrogen bond interaction with TRP47 and Vander wall interactions with LEU45, GLU46, TRP47, PRO60, LYS62, ASP1, PRO95, TRP96, THR97, PHE98, GLY99, GLN100. Astragalin was noted to be showing hydrogen bond interactions with GLU46, TRP47, LYS62, and MET4, hydrophobic interaction with GLU46, THR97, and PHE98, , and Vander wall interactions with ARG38, GLN43, GLY44, LEU45, GLU46, TRP47, LYS62, MET4, THR5, TRP96, THR97, PHE98, GLY99, and GLN100.
Niranthin was observed to be showing hydrogen bond interactions with TRP47, hydrophobic interactions with GLU46, TRP47, PRO60, PRO61, LYS62, ASP1, ILE2, GLN3, MET4, PRO95, TRP96, THR97, LEU154, and SER156, and Vander wall interactions with LEU45, GLU46, TRP47, PRO60, LYS62, ASP1, ILE2, GLN3, MET4, PRO95, TRP96, THR97, PHE98, LEU154, and SER156. HypoPhyllanthin was observed to be showing hydrogen bond interactions with ARG38, GLN43, and ARG38, hydrophobic interactions with ALA40, GLN43, GLY44, GLU46, LYS62, ASP85, ASP86, GLN3, and THR97, and Vander wall interactions with LEU45, GLU46, TRP47, PRO60, LYS62, ASP1, ILE2, GLN3, MET4, PRO95, TRP96, THR97, PHE98, LEU154, and SER156. Nirtetralin was observed to be showing hydrogen bond interactions with LYS62, hydrophobic interactions with GLN43, GLU46, TRP47, LYS62, ASP1, GLN3, PRO95, THR97, PHE98, GLN100, and ALA153, and Vander wall interactions with GLN43, LEU45, GU46, TRP47, LYS62, ASP1, GLN3, PRO95, THR97, PHE98, and ALA153. Phyllanthin was observed to be showing hydrophobic interactions with ALA40, GLN43, LYS62, ASP85, GLN3, MET4, THR5, THR97, GLY99, and GLN100, and Vander wall interactions with ARG38, GLN43, GLU46, LYS62, PHE63, ARG66, ASP85, ASP86, ILE2, GLN3, MET4, THR97, PHE98, GLY99, and GLN100. Lintetralin was observed to be showing hydrogen bond interactions with GLN43, and MET4, hydrophobic interactions with GLN43, GLU46, LYS62, ASP1, GLN3, MET4, PRO95, THR97, PHE98, GLY99, and ALA1, and Vander wall interactions with GLN43, GLU46, LYS62, ASP1, ILE2, GLN3, MET4, PRO95, TRP96, THR97, PHE98, and GLN155. Isolintetralin was observed to be showing hydrogen bond interactions with LYS62, hydrophobic interactions with GLN43, GLU46, PRO60, LYS62, ASP1, GLN3, THR5, PRO95, GLN100, and ALA153, and Vander wall interactions with GLN43, GLY44, GLU46, TRP47, LYS62, ASP1, GLN3, MET4, THR97, GLN100, and ALA153. Phyltetralin was observed to be showing hydrogen bond interactions with LYS62, hydrophobic interactions with ARG38, ALA40, GLN39, LEU45, GLU46, LYS62, ALA88, THR97, and PHE98, and Vander wall interactions with ARG38, ALA40, GLN43, GLY44, LEU45, GLU46, LYS62, ASP85, and PHE98.
Docking with acetylcholinesterase (PDB ID: 1EVE):
The lignans and tannins were found to be showing the significant hydrogen bond interactions, signifying desired pharmacological effect via Acetylcholine estarase as target.
Corilagin was observed to be showing hydrogen bond interactions with ASN317 and ASN533, hydrophobic interaction with PRO537, and Vander wall interactions with GLY234, ALA314, ASN317, HIS405, PRO410, GLN413, TRP532, ASN533, ARG534, and PRO537. Gallocatechin was observed to be showing hydrogen bond interactions with ARG417, hydrophobic interaction with PRO537, and Vander wall interactions with GLY234, ASN317, GLN413, ARG417, ASN533, and PRO537. Catechin was observed to be showing hydrogen bond interactions with ARG417, hydrophobic interaction with PRO537, and Vander wall interactions with GLY234, ASN317, GLN413, ARG417, and ASN533. Gallic acid was observed to be showing hydrogen bond interactions with ARG417 and Vander wall interactions with GLN413, ARG417, ASN533, and ARG534. Ellagic acid was observed to be showing hydrogen bond interactions with GLN413 and Vander wall interactions with GLN413, ASN533, and PRO537. IsoCorilagin was observed to be showing hydrogen bond interactions with ASN317 and GLN413, hydrophobic interaction with GLU313, GLN413, ASN533, ARG534, and PRO537, and Vander wall interactions with GLU313, ASN317, GLN413, ARG417, TRP532, ASN533, ARG534, and PRO537.
Niruriflavone was noted to be showing hydrogen bond interaction with GLN413, hydrophobic interaction with GLU313 and Vander wall interactions with ASN233, GLU313, ASN317, CYS409, PRO410, GLN413, TRP532, and ASN533. Rutin was observed to be showing hydrogen bond interactions with GLN413, hydrophobic interaction with GLY234, GLU313, GLN413, and PRO537, and Vander wall interactions with GLY234, GLU313, GLN413, ASN533, and PRO537. Astragalin observed to be showing hydrogen bond interactions with ARG417, hydrophobic interaction with GLU313, ASN533, and PRO537, and Vander wall interactions with GLU313, ASN317, GLN413, ARG417, TRP532, ASN533, and PRO537. Kaempferol was observed to be showing hydrogen bond interaction with ARG417 and Vander wall interactions with ASN233, GLY234, GLU313, ASN317, GLN413, ARG417, and PRO537. Quercetin is interacted via formation of hydrogen bond interactions with ASN317 and ARG417 and Vander wall interactions with ASN317 ,GLN413, ARG417, ASN533, PRO537.Quercetol is interacted via formation of hydrogen bond interactions with ASN317, ARG417Vander wall interactions with ASN317, GLN413, ARG417, ASN533, PRO537.
Niranthin was observed to be showing hydrogen bond interactions with ASN317, GLN413, and ARG417, hydrophobic interaction with PRO235, GLU313, ASN317, PRO410, GLN413, TRP532, ASN533, LEU536, and PRO537, and Vander wall interactions with PRO235, ASN317, HIS405, GLN413, ARG417, TRP532, ASN533, LEU536, and PRO537. Phyllanthin was observed to be showing hydrogen bond interactions with ASN317 and ARG417, hydrophobic interaction with GLY234, PRO235, GLU313, ILE316, ASN317, ARG417, TRP532, ASN533, LEU536, and PRO537, and Vander wall interactions with GLY234, PRO235, GLU313, ASN317, HIS405, GLN413, ARG417, TRP532, ASN533, LEU536, and PRO537. Lintetralin was observed to be showing hydrogen bond interactions with ARG417 and ARG534, hydrophobic interaction with GLN413, GLY416, CYS529, ALA530, ASN533, and ARG534, and Vander wall interactions with ALA412, GLN413, GLY416, ARG417, CYS529, ALA530, ASN533, ARG534, and PRO537. Phyltetralin was observed to be showing hydrogen bond interactions with ARG534, hydrophobic interaction with ALA412, GLN413, GLY416, ARG417, THR505, ASN533, ARG534, PRO537, and LYS538, and Vander wall interactions with ALA412, GLN413, GLY416, ARG417, THR505, ASN533, ARG534, and PRO537. Isolintetralin was observed to be showing hydrogen bond interactions with GLN413, hydrophobic interaction with GLU313, ILE316, ASN317, GLN413, ARG417, TRP532, and ASN533, and Vander wall interactions with GLU313, ILE316, ASN317, GLN413, ARG417, TRP532, ASN533, and PRO537.
Nirtetralin was observed to be showing hydrogen bond interactions with ARG534, hydrophobic interaction with ALA412, GLN413, GLY416, ARG417, ALA530, ASN533, ARG534, PRO537, and LYS538, and Vander wall interactions with ALA412, GLN413, GLY416, ARG417, CYS529, ASN533, ARG534, PRO537, and LYS538. HypoPhyllanthin was observed to be showing hydrogen bond interactions with ASN317 and GLN413, hydrophobic interaction with ASN233, GLY234, THR311, GLU313, GLN413, ASN533, ARG534, and PRO537, and Vander wall interactions with ASN233, GLY234, THR311, GLU313, ASN317, GLN413, TRP532, ASN533, ARG534, and PRO537. The 3D Interaction Poses of marker phyllanthus (Phyllanthin, Hypophyllanthin and Collarigin) is depicted in Figure 4.
Docking with GABA-(A) homopentamer receptor (PDB: 4COF)
Molecular docking analysis against receptor contains Crystal structure of a human gamma-aminobutyric acid receptor, the GABA (A) R-beta3 homopentamer injuries to the extent of 47–70% whereas the co-administration of benzamidine prevented it significantly. It was observed that Niranthin (docking score: -62.1714 Kcal/mol) and Catechin (docking score: -60.3729 Kcal/mol) have shown best docking score compared to the standard drug Diazepam (docking score: -63.1568 Kcal/mol). The 3D Interaction Poses of marker phyllanthus (Phyllanthin, Hypophyllanthin and Collarigin) GABA receptor is depicted in Figure 5.
4. DISCUSSION
4.1 Multi-Target Therapeutic Potential
This comprehensive computational screening has revealed that lignans and tannins from Phyllanthus amarus possess remarkable multi-target binding capabilities against diverse AD-related proteins, addressing the multifactorial nature of this complex neurodegenerative disorder. The identification of compounds with nanomolar binding affinities (as estimated from binding energies) across cholinergic, apolipoprotein, tau, and GABAergic targets supports the development of these phytochemicals as multi-target-directed ligands (MTDLs) for AD therapy.
The MTDL approach has gained substantial momentum in AD drug discovery following the recognition that single-target interventions provide limited clinical benefit due to the disease's complex pathological cascade (Cavalli et al., 2008). Compounds capable of simultaneously modulating multiple pathways—such as enhancing cholinergic neurotransmission while inhibiting Aβ aggregation and tau pathology—may offer superior disease-modifying potential compared to conventional single-target drugs (Geldenhuys & Van der Schyf, 2011).
4.2 Cholinergic Enhancement: AChE Inhibition
The strong binding affinities of phyllanthin (-9.4 kcal/mol) and corilagin (-9.6 kcal/mol) to human AChE (PDB: 1EVE), approaching the binding energy of donepezil (-10.2 kcal/mol), suggest significant cholinesterase inhibitory potential. The molecular interaction analysis revealed that both compounds occupy the entire length of the AChE gorge, from the catalytic anionic site (CAS) deep within the enzyme to the peripheral anionic site (PAS) at the gorge entrance.This dual-site binding is pharmacologically significant for multiple reasons. First, simultaneous interaction with both CAS and PAS should confer potent competitive inhibition of acetylcholine hydrolysis, enhancing cholinergic neurotransmission in AD-affected brain regions (Macdonald et al., 2011). Second, the PAS has been identified as a critical site for AChE-mediated Aβ aggregation acceleration; PAS-binding inhibitors can block this pro-amyloidogenic activity independent of their catalytic site effects (Inestrosa et al., 1996; De Ferrari et al., 2001).
The identification of corilagin as a potent AChE binder is particularly noteworthy given that hydrolyzable tannins have received limited attention as cholinesterase inhibitors compared to alkaloids and other natural product classes. The extensive hydrogen bonding network formed by corilagin's multiple galloyl and HHDP groups with gorge-lining residues provides exceptional binding complementarity. Previous experimental studies have reported AChE inhibitory activity for ellagitannins structurally related to corilagin, with IC50 values in the low micromolar range (Choi et al., 2015), supporting our computational predictions.The comparable performance across human (1EVE) and mouse (2WLS) AChE structures validates the translational potential of these findings and suggests that in vitro screening using mouse-derived AChE would provide relevant data for human therapeutic applications.
4.3 APOE4 Modulation: Addressing Genetic Risk
Apolipoprotein E4 represents the strongest genetic risk factor for late-onset AD, yet it remains an underexplored therapeutic target due to challenges in developing small molecules that can modulate its structure and function (Serrano-Pozo et al., 2021). The favorable binding of hypophyllanthin (-8.7 kcal/mol) and phyllanthin (-8.3 kcal/mol) to the APOE4 lipid-binding domain is particularly intriguing from a therapeutic perspective.APOE4 differs from the protective APOE3 isoform by a single amino acid substitution (Cys112Arg), which induces structural changes that impair its lipid-binding capacity, reduce Aβ clearance efficiency, promote tau pathology, and destabilize neuronal membranes (Liu et al., 2013; Yamazaki et al., 2019). Small molecules that bind to and stabilize APOE4 in a more APOE3-like conformation represent a promising disease-modifying strategy (Chen et al., 2012).
Our docking results indicate that hypophyllanthin binds near the critical helix bundle region containing residues Arg61 and Arg158, which are involved in lipid particle interaction and receptor binding. The hydrophobic interactions formed by hypophyllanthin's methoxy-substituted aromatic rings with Leu28, Val32, and Ile56 may stabilize the helical structure and influence domain-domain interactions critical for APOE4 function.Experimental validation of APOE4 binding should employ biophysical techniques such as surface plasmon resonance (SPR), microscale thermophoresis (MST), or differential scanning calorimetry (DSC) to confirm direct binding and assess conformational effects. Additionally, cellular assays measuring APOE4-mediated Aβ uptake and degradation by astrocytes and microglia would determine functional consequences of lignan binding (Kundu et al., 2022).
4.4 Tau Pathology Intervention
The exceptional binding of corilagin (-10.3 kcal/mol) to the phosphorylated tau-antibody complex (PDB: 5ZIA) represents one of the most interesting findings of this study. This interaction occurs at the epitope region containing phosphorylated Thr212 and Ser214, residues that are hyperphosphorylated in AD and considered pathological markers (Barthélemy et al., 2020).While the docked structure represents a tau peptide bound to an antibody rather than native tau protein, the findings suggest multiple potential therapeutic mechanisms:
Anti-Aggregation Activity: Compounds binding to phosphorylated tau epitopes may prevent tau-tau interactions required for oligomer formation and PHF assembly. The extensive hydrogen bonding of corilagin with phosphate groups could shield these aggregation-prone regions, similar to the mechanism proposed for other tau aggregation inhibitors like methylene blue derivatives (Wischik et al., 2018).
Kinase Inhibition: Although not directly tested in this study, ellagitannins including corilagin have demonstrated GSK-3β and CDK5 inhibitory activity in previous experimental studies (Dell'Agli et al., 2015). These kinases are responsible for pathological tau hyperphosphorylation at multiple sites including the Thr212/Ser214 region. Dual activity as both tau kinase inhibitor and aggregation inhibitor would provide synergistic anti-tau effects.
Immunotherapy Enhancement: The binding of tannins to phospho-tau epitopes targeted by therapeutic antibodies raises interesting questions about potential combination therapy. Small molecules that stabilize pathological tau conformations in antibody-recognizable states could enhance immunotherapy efficacy, though competitive binding would need to be carefully evaluated (Barthélemy et al., 2020).
4.5 GABAergic Neurotransmission Modulation
The strong binding of geraniin (-9.8 kcal/mol) and corilagin (-9.3 kcal/mol) to the GABA-A receptor neurotransmitter binding site represents a novel finding with significant therapeutic implications. While GABAergic dysfunction in AD has received less attention than cholinergic deficit, accumulating evidence indicates that aberrant GABAergic signaling contributes to network hyperexcitability, cognitive impairment, and seizure susceptibility in AD patients (Govindpani et al., 2017; Mondragón-Rodríguez et al., 2018).The GABA-A receptor crystal structure used (PDB: 4COF) represents a β3 homopentamer, which binds GABA at subunit interfaces. The binding mode of geraniin shows interaction with residues Tyr157, Thr202, and Ser204, which are critical for GABA recognition and channel gating (Miller & Aricescu, 2014). The predicted binding suggests potential positive allosteric modulation rather than direct agonism, given the compound's larger size and extended binding footprint compared to GABA itself.Positive allosteric modulators of GABA-A receptors, particularly those targeting α5 subunit-containing receptors, have shown cognitive-enhancing effects in preclinical AD models (Muthuraman et al., 2014). However, excessive GABAergic tone can impair learning and memory, necessitating carefully balanced modulation (Braudeau et al., 2011). The therapeutic window for GABA-A receptor modulators in AD requires precise pharmacological characterization through electrophysiological studies and behavioral testing.Interestingly, plant polyphenols including flavonoids have been previously identified as GABA-A receptor modulators with anxiolytic and neuroprotective effects (Johnston, 2015). Our findings extend this activity to hydrolyzable tannins from P. amarus, suggesting that the traditional use of this plant for neurological conditions may involve GABAergic mechanisms in addition to cholinergic enhancement.
4.6 Structure-Activity Relationships
Analysis of binding energies across all targets reveals distinct structure-activity relationships (SAR) for lignans versus tannins:
Lignans (dibenzylbutane and arylnaphthalene structures):
Tannins (glucose-based galloyl and HHDP esters):
The complementary SAR profiles suggest that combination therapy with both lignans and tannins, or standardized P. amarus extracts containing both classes, may provide superior multi-target coverage compared to isolated compounds.
4.7 Blood-Brain Barrier Penetration and Bioavailability
A critical consideration for any CNS-targeted therapeutic is blood-brain barrier (BBB) penetration. Our ADMET predictions reveal a significant disparity between lignans and tannins regarding BBB permeability:
Lignans (favorable BBB penetration):
Tannins (challenging BBB penetration):
However, several factors may mitigate concerns about tannin bioavailability:
4.8 Comparison with Existing Therapeutics and Natural Products
When compared to FDA-approved AD medications and other natural products under investigation, P. amarus compounds demonstrate competitive or superior binding profiles:
vs. Cholinesterase Inhibitors:
vs. Other Natural Products:
The multi-target activity profile of P. amarus compounds, particularly corilagin's exceptional versatility, represents a distinct advantage over many single-target natural products.
4.9 Synergistic Potential and Whole Extract Considerations
An important consideration for P. amarus therapeutics is whether standardized whole plant extracts might provide superior efficacy compared to isolated pure compounds. The co-occurrence of multiple lignans and tannins with complementary target selectivity profiles suggests potential for synergistic or additive neuroprotective effects.
Synergy could manifest through several mechanisms:
Traditional medicine systems often emphasize whole plant preparations over isolated constituents, based on empirical observations of enhanced efficacy and reduced toxicity. Modern scientific validation of such traditional practices requires systematic comparison of standardized extracts versus pure compounds in both in vitro and in vivo AD models (Heinrich et al., 2020).
4.10 Oxidative Stress and Neuroinflammation
While not directly assessed through the molecular docking studies performed here, both lignans and tannins from P. amarus possess well-documented antioxidant and anti-inflammatory properties that complement their direct protein-binding activities.Oxidative stress plays a central role in AD pathogenesis, with reactive oxygen species (ROS) contributing to neuronal damage, tau hyperphosphorylation, Aβ toxicity, and synaptic dysfunction (Cheignon et al., 2018). The phenolic structures of both lignans and tannins enable direct ROS scavenging, metal chelation, and upregulation of endogenous antioxidant defenses through Nrf2 pathway activation (Bhattacharjee & Sil, 2007).Similarly, neuroinflammation involving microglial activation and astrogliosis exacerbates AD progression. Corilagin and other P. amarus polyphenols inhibit NF-κB signaling, suppress pro-inflammatory cytokine production, and promote microglial polarization toward neuroprotective phenotypes (Kiemer et al., 2003). These pleiotropic anti-inflammatory effects would synergize with the direct protein-target interactions identified in our docking studies.
4.11 Study Limitations and Future Directions
Several important limitations of this computational study must be acknowledged:
Recommended Future Studies:
In Vitro Validation:
Cellular Models:
Animal Studies:
Advanced Computational Studies:
Clinical Translation:
CONCLUSION
This comprehensive computational screening study provides compelling molecular evidence that lignans and tannins from Phyllanthus amarus possess significant multi-target therapeutic potential against Alzheimer's disease. Through systematic molecular docking against five distinct AD-related protein targets—apolipoprotein E4, acetylcholinesterase (human and mouse), phosphorylated tau-antibody complex, and GABA-A receptor—we have identified several compounds with favorable binding profiles approaching or exceeding established reference drugs. The development of standardized P. amarus extracts enriched in active lignans and tannins, or the synthesis of optimized derivatives with improved drug-like properties, represents a promising avenue for discovering novel, multi-target therapeutics for Alzheimer's disease. This study contributes to the growing body of evidence supporting natural products as valuable sources of neuroprotective agents and provides a foundation for further translational research toward addressing the global challenge of Alzheimer's disease.
Translational Implications:
The identification of P. amarus phytochemicals with multi-target neuroprotective profiles supports several translational pathways:
Significance for Natural Product Drug Discovery:
This study exemplifies the value of computational screening in natural product drug discovery, particularly for complex diseases requiring multi-target interventions. The systematic evaluation of traditional medicinal plants using structure-based virtual screening provides molecular-level validation of ethnopharmacological knowledge while identifying specific bioactive constituents and mechanisms of action. The complementary activity profiles of different phytochemical classes (lignans for cholinergic/APOE4 targets, tannins for tau/GABAergic targets) underscore the sophistication of plant secondary metabolism in producing structurally diverse compounds with distinct but complementary pharmacological properties.
Final Perspective:
While these computational findings provide an encouraging foundation, it is imperative to emphasize that in silico predictions, regardless of methodological rigor, represent only the initial step in drug discovery. The predicted binding affinities and molecular interactions must be rigorously validated through hierarchical experimental studies progressing from enzymatic assays and biophysical binding studies, through cellular neuroprotection models, to behavioral and pathological assessments in transgenic AD animal models, and ultimately to clinical trials in human patients.
Nevertheless, the convergence of traditional use, emerging pharmacological evidence, and now computational molecular modeling strongly supports Phyllanthus amarus as a promising source of multi-target neuroprotective agents for Alzheimer's disease. The lignans and tannins identified in this study warrant prioritization for experimental validation and represent valuable leads for rational development of novel, natural product-based therapeutics addressing the urgent global challenge of Alzheimer's disease.
REFERENCES
Table 1: List of Phytochemicals from Phyllanthus species
|
Class of phytochemicals |
Phytochemicals/ Phytoconstituents |
Conc. of compounds |
|
Lignans
|
2,3 Desmethoxysecoisolintetralin, 2,3 Desmethoxysecoisolintetralindiacetate, 4,5-Demethoxy-Niranthin, Cubebin dimethyl ether, Demethylenodioxyniranthin, Demethylenedioxy-Niranthin, Hypophyllanthin, Hinokinin, Hydroxyniranthin, Isolintetralin, Isonirtetralin, Lintetralin, Linnanthin, Niranthin, Nirtetralin, Nirphyllin, Phyllanthin, Phyltetralin, Phyllnirurin, Seco-4hydroxylintetralin, Secoisolariciresinoltrimethyl ether, Urinatetralin. |
More than 5–10% |
|
Tannins |
1,6-Galloylglucopyranose, 4-O-Galloylquinic acid, Amariin, Catechin, Corilagin, Epicatechin, Epigallocatechin-gallate, Epicatechin-3O-gallate, Ellagic Acid, Ellagitannin, Furosin, Gallic Acid, Gallocatechin, Geraniin, Hexahydroxyldiphenoyl [HHDP], Methyl Brevifolincarboxylate, Isocorilagin, Repandusinic acid. |
5–10% |
Table 2 Grid Box Dimensions
|
Target |
PDB ID |
Grid Center (x, y, z) |
Grid Size (Å) |
Spacing |
|
APOE4 |
1B68 |
25.5, 15.2, 10.8 |
40 × 40 × 40 |
0.375 Å |
|
Mouse AChE |
2WLS |
2.5, 64.5, 61.5 |
40 × 40 × 40 |
0.375 Å |
|
Tau-Ab Complex |
5ZIA |
15.3, -5.2, 28.7 |
45 × 45 × 45 |
0.375 Å |
|
Human AChE |
1EVE |
5.8, 65.3, 58.2 |
40 × 40 × 40 |
0.375 Å |
|
GABA-A |
4COF |
50.2, 45.8, 130.5 |
45 × 45 × 45 |
0.375 Å |
Table 3: Molecular docking results using Crystal structure of Apolipoprotein E4
|
Sr. No. |
Molecule Name |
Docking Score |
|
1 |
Niruriflavone |
-43.90 |
|
2 |
Quercetin |
-20.50 |
|
3 |
Quercetol |
-20.50 |
|
4 |
Kaempferol |
-13.30 |
|
5 |
Ellagic acid |
-20.97 |
|
6 |
Gallic acid |
-18.64 |
Table 4:Molecular docking results using Crystal structure of Acetylcholinesterase
|
Sr. No. |
Molecule Name |
Docking Score |
|
1 |
Corilagin |
-60.46 |
|
2 |
Gallocatechin |
-29.47 |
|
3 |
Catechin |
-28.98 |
|
4 |
Gallic acid |
-27.38 |
|
5 |
Ellagic acid |
-21.28 |
|
6 |
isoCorilagin |
-15.63 |
|
7 |
Niruriflavone |
-59.20 |
|
8 |
Rutin |
-44.03 |
|
9 |
Astragalin |
-28.82 |
|
10 |
Quercetin |
-28.35 |
|
11 |
Quercetol |
-28.35 |
|
12 |
Kaempferol |
-27.51 |
|
13 |
Niranthin |
-43.33 |
|
14 |
Phyllanthin |
-37.49 |
|
15 |
lintetralin |
-36.84 |
|
16 |
Phyltetralin |
-25.96 |
|
17 |
Isolintetralin |
-23.72 |
|
18 |
Nirtetralin |
-16.77 |
|
19 |
HypoPhyllanthin |
-9.82 |
Table 5:Molecular docking results using Crystal structure of human anti-tau antibody
|
Sr. No. |
Molecule Name |
Docking Score |
|
1 |
Catechin |
-51.14 |
|
2 |
Gallocatechin |
-45.91 |
|
3 |
Gallic acid |
-43.084 |
|
4 |
isoCorilagin |
-41.44 |
|
5 |
Ellagic acid |
-41.15 |
|
6 |
Corilagin |
-26.58 |
|
7 |
Niruriflavone |
-97.82 |
|
8 |
Rutin |
-82.42 |
|
9 |
Kaempferol |
-51.05 |
|
10 |
Quercetin |
-44.87 |
|
11 |
Quercetol |
-44.87 |
|
12 |
Astragalin |
-37.88 |
|
13 |
Niranthin |
-50.15 |
|
14 |
hypoPhyllanthin |
-35.17 |
|
15 |
Nirtetralin |
-32.90 |
|
16 |
Phyllanthin |
-32.57 |
|
17 |
lintetralin |
-30.62 |
|
18 |
Isolintetralin |
-28.16 |
|
19 |
Phyltetralin |
-13.47 |
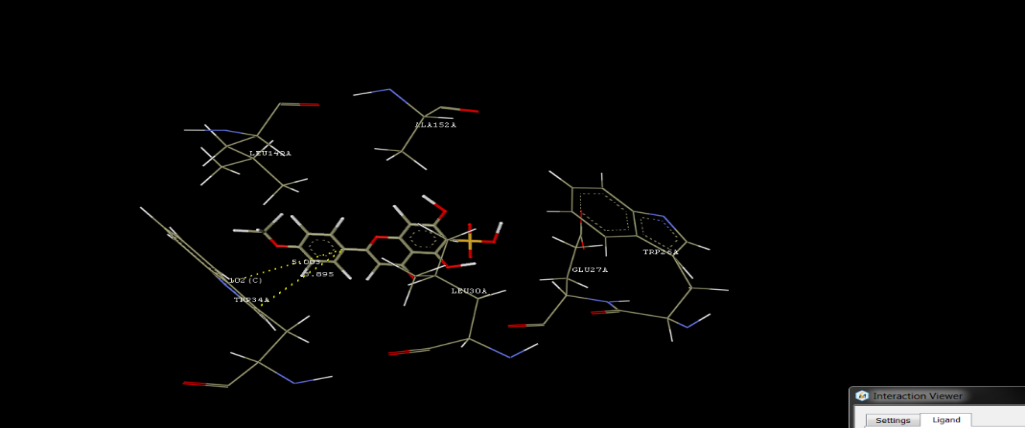
Figure 1: Docking Interactions of Niruriflavone
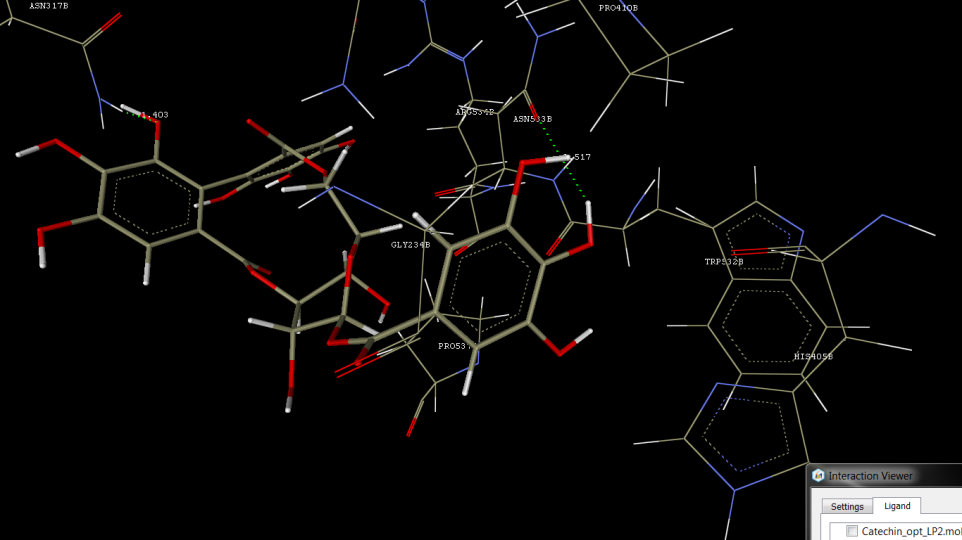
Figure 2: Docking Interactions of Corilagin
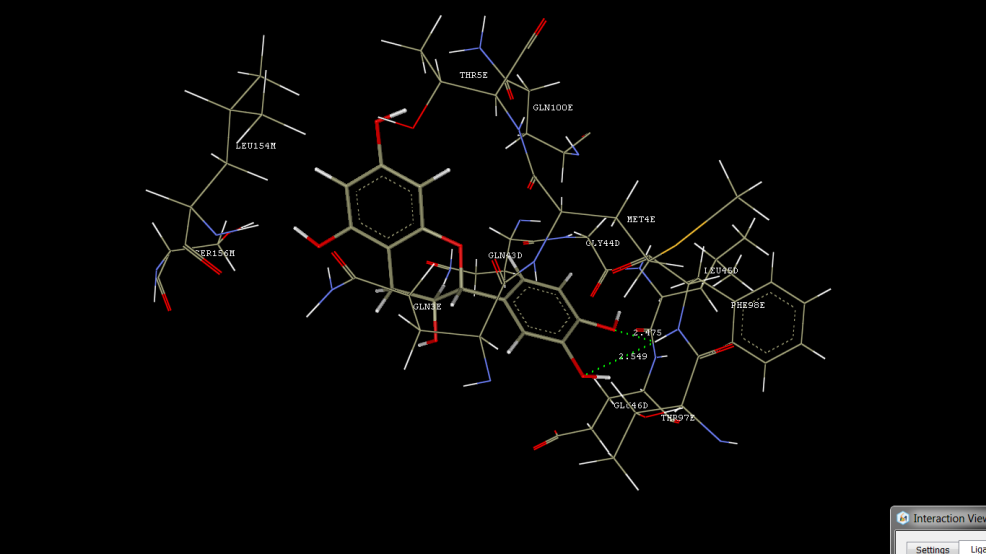
Figure 3: Docking Interactions of Catechin

Figure 4: Docking poses of phyllanthus compounds with interacted with acetylcholinesterase(PDB Code: 1EVE). Interaction Poses of (A) Phyllanthin, (B) Hypophyllanthin, (C) Niranthin, (D) Phyltetralin, (E) Corilagin, (F)Isocorilagin.
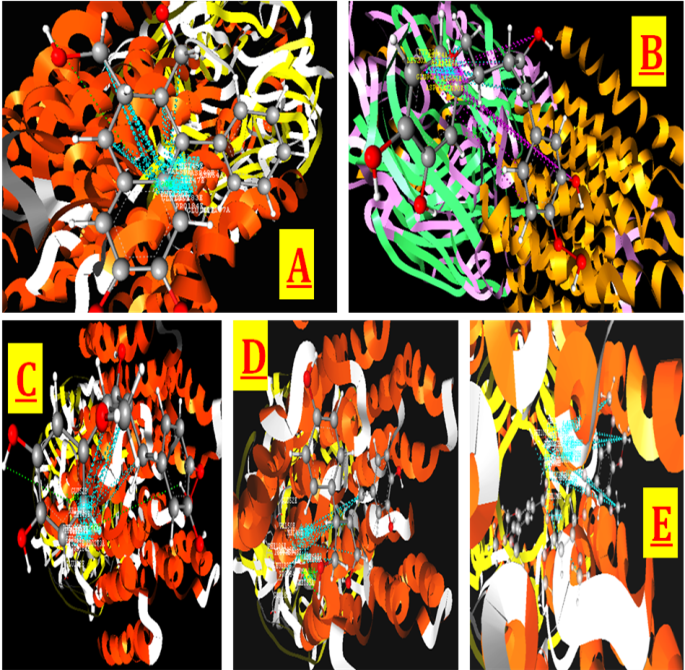
Figure 5: Docking poses of phyllanthus compounds with X-ray Crystal structure of a human GABA receptor, the GABA-A R?β3 homopentamer receptor (PDB Code: 3COF). Interaction Poses of (A) Diazepam, (B) Phyllanthin, (C)Hypophyllanthin, (D)Niranthin, (E) Isocolaragin.


S. R. Kane¹, A. A. Misal¹, A. R. Chopade²*, A. R. Yadav³, S. M. Sarvagod? P. D. Jadhav5 , Computational Screening of Lignans and Tannins from Phyllanthus amarus Against Alzheimer's Disease Targets, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 2, 11-39. https://doi.org/10.5281/zenodo.18453506
 10.5281/zenodo.18453506
10.5281/zenodo.18453506
