1 Global College of Pharmaceutical Technology, Krishnanagar-741102, West Bengal, India
2 Karimpur Pannadevi College, Karimpur-741152, West Bengal, India
Resistance developing in some of the world's most common bacteria is a major concern for public health, according to experts throughout the world. Since multidrug-resistant organisms have emerged in both healthcare facilities and general public, it is reasonable to assume that antibiotic-resistant bacterial reservoirs exist outside of hospitals as well. Bacterial adaptation to antibiotic "attack" is the pinnacle of evolution and bacterial adaptability. The "survival of the fittest" is the result of bacterial pathogens' remarkable genetic plasticity, which can evolve resistance to nearly all antibiotics used in clinical practice through mutational adaptations, gene acquisition, or changes in gene expression. Consequently, in order to combat organisms that are resistant to several medications and to stop the development and spread of resistance, it is essential to understand the physiological and genetic bases of resistance. This study will focus on the primary mechanisms of antibiotic resistance that manifest in clinical practice, with specific instances drawn from relevant bacterial infections. A major new cause of death and sickness on a worldwide scale is the development of resistance to antimicrobial agents. Upon their first introduction in the 1900s, antibiotics were believed to have triumphed against bacteria. Unfortunately, it didn't take long for researchers to figure out that bacteria may develop resistance to any medicine. It seems that even certain antimicrobial therapies may be developed into resistance by the bulk of harmful bacteria. Inactivation of the drug, modification of the drug's target, restriction of drug uptake, and active efflux are the four main resistance mechanisms. The microorganisms could have these abilities either by birth or by microbial transmission. Better treatment options for infectious diseases and antimicrobial drugs that withstand microbial attempts to build resistance may result from our growing understanding of these pathways.
Antibiotic resistance has been around for a while, and the "resistome" is an ever-changing and expanding threat. Many causes contribute to the worldwide resistance, such as overpopulation, increasing migration throughout the world, antibiotic usage in animal production and healthcare, natural selection, lack of sanitation, wildlife expansion, and insufficient sewage disposal systems (Singer et al., 2016). The use of antibiotics is one of the main ways that modern medicine treats infections. Many antibiotics were created between the 1930s and the 1960s, a period known as the "golden era" of antibiotics (Nathan and Cars, 2014). The sad end to this era came when researchers had to scale down the pace of new antibiotic discoveries due to microbes that were becoming more resistant to existing ones. Careless antibiotic use and the ongoing inability to create or discover novel medications are risk factors for the emergence of antibiotic resistance (Nathan, 2004). Antimicrobial resistance (AMR) is a growing problem that threatens the health of humans, animals, and the environment across the world. The rise, spread, and persistence of "superbugs," or bacteria resistant to a wide range of medications, is to blame (Davies and Davies, 2010). There is a two-way flow of infectious diseases within the animal, human, and environmental triangle, which includes multidrug-resistant (MDR) bacteria. Some of the probable reasons for "the global resistome," or AMR, include the overuse of antibiotics in both animals and humans (in food, pets, and aquatic environments), the sale of antibiotics without a prescription, an increase in international travel, poor sanitation and hygiene, and the discharge of nonmetabolized antibiotics or their residues into the environment through manure or feces (Fig. 1). Multiple factors contribute to the genetic selection pressure that affects the prevalence of multidrug-resistant microorganisms in a population. Recent data on antibiotic use in cattle has shown continent-specific antibiotic hotspots, which might have long-term effects on public health and the economy. Predictions indicate that the usage of antibiotics in livestock, including pigs, poultry, and cattle, will reach 67% in the world's most populated countries by 2030 (Van Boeckel et al., 2015). Antibacterial agents lose some of their effectiveness when tolerance or resistance develops even after a single application of this chemical. This class of medications includes antimicrobials that combat infections caused by bacteria, viruses, fungi, and parasites. This newfound resistance could be guided by a myriad of biochemical and physiological mechanisms. The establishment and expansion of the resistance required several complicated processes, which must not be exaggerated. Further, a major obstacle that has halted several significant advances in the management of resistance development is the paucity of fundamental data on these specific subjects. This is a major global public health concern, and many groups and organizations have recognized it. Little has changed thus far, despite the many resolutions, studies, and ideas that have been made. Unfortunately, antibiotic resistance is on the rise again (Roca et al., 2015).

Figure 1: Different kinds of factors/causes that lead to antibiotic resistance.
Historical Background of antibiotics:
In the late 1950s and early 1960s, it was first discovered that enteric bacteria, such as Shigella, Escherichia coli, and Salmonella, could withstand several antimicrobial treatments. Clinical, financial, and human losses were substantial as a consequence of these resistant strains, which mostly affected impoverished nations. The industrialized world, however, dismissed it as a small health concern affecting only gut microbes. The discovery of ampicillin-resistant strains of Neisseria gonorrhoeae and Haemophilus influenzae, as well as Haemophilus's resistance to tetracycline and chloramphenicol, put an end to this theory in the 1970s. The increasing use of antimicrobials, particularly in developing countries where they were easily accessible over-the-counter, led to a rise in the incidence of resistance. New and powerful antibiotics were hard to come by due to a lack of funding for medical care and unsanitary living circumstances, both of which facilitated the emergence of resistance (Rossolini et al., 2014). Many prominent groups have recognized antibiotic resistance as a "global public health concern". These groups include the World Economic Forum, the Infectious Diseases Society of America (IDSA), the Centers for Disease Control and Prevention (CDC), and the World Health Organization (WHO). In order to address the problem of antibiotic resistance, the World Health Assembly requested WHO to develop a global strategy (Payne et al., 2015). Globally, antibiotic resistance is not decreasing; rather, it may be trending in an opposite way. Worldwide, people are feeling the impacts of antibiotic resistance, which may have several origins. Many aspects of antibiotic resistance have been the subject of much research, and many possible solutions to this global problem have been tried and evaluated. In particular on a worldwide political scale, none of them are particularly well-organized (Prestinaci et al., 2015). Antibiotics are now an essential part of every healthcare system due to their revolutionary effect on both medical and social advancement. In the absence of effective antibiotic therapy to prevent bacterial infections, the achievements of modern medicine, which include complex major surgeries, cancer treatment, organ transplants, and care for preterm babies, could not have been possible. To save future social, medical, and economic opportunities from disaster, effective worldwide action plans must be adopted. Our goal here is to show the global size, key causes, and the impacts, drawing attention to critical areas that need swift action (Laxminarayan et al., 2013).
Antibiotic discovery and supremacy:
Records of microbial disease management date back to many different parts of the globe, from China to Greece. The discovery of penicillin in 1928 ushered in the modern era of medicine (Sengupta et al., 2013). Countless lives have been spared, and medical history has altered because of these "magic bullets" since then. In the 1940s, the first prescription for an antibiotic was issued. For bacterial illnesses, penicillin was the drug of choice in the armed forces during WWII (Piddock, 2012). Unfortunately, penicillin resistance quickly became a major clinical concern. Therefore, much of the progress made in the 1940s was in jeopardy during the 1950s. However, scientists wasted little time in discovering and developing new β-lactam antibiotics so that antibiotic treatment schedules could be maintained. The United Kingdom and the United States both recorded the first instances of methicillin-resistant Staphylococcus aureus (MRSA) in the same decade, 1962 and 1968, respectively, when new medications were introduced (Spellberg and Gilbert, 2014). A number of medical and surgical advancements owe a debt of gratitude to antibiotics, which have been instrumental in combating the worldwide problem of antibiotic resistance. Antibiotics have been used to treat or prevent bacterial infections that may develop after chemotherapy or major operations like organ transplants, hip replacements, or heart surgeries. These miraculous treatments have effectively treated bacterial infections, resulting in a rise in the average life expectancy. In 1920, Americans could expect to live an average of 56.4 years; now, that figure hovers around 80 years (Rossolini et al., 2014). There are other regions of the globe where antibiotics have been useful; for example, in developing countries with weak public health systems, they have reduced the number of cases of bacterial infections in livestock and people (Wright, 2014).
Superbugs and super-resistance:
"Superbugs" are bacteria that have mutated to become resistant to many types of antibiotics, making them more dangerous and potentially fatal. Longer hospital stays and increased expenses are associated with these antibiotic-resistant microbes, and there are fewer treatment options for them. At times, super resistant strains have shown signs of becoming more pathogenic and deadly (Davies and Davies, 2010). Many bacterial human illnesses have evolved into multidrug-resistant strains as a result of antibiotic overuse. One well-known example of a disease that may emerge in both developed and developing countries is multidrug-resistant tuberculosis (Shah et al., 2007). Acinetobacter, Burkholderia, Campylobacter, Citrobacter, clostridium, Enterobacter spp., Haemophilus, Klebsiella, Proteus, Salmonella, Serratia, Staphylococcus aureus, and Staphylococcus epidermidis are among the other dangerous bacteria that may cause infections (Sotgiu et al., 2009). S. aureus is often believed to be the most notorious superbug. It causes skin infections and is a frequent human nasal commensal. A major public health concern nowadays is CA-MRSA with increased acquired virulence (DeLeo and Chambers, 2009). Different mec gene clusters and genes encoding the cytotoxic leukocidin Panton-Valentine are two examples of the distinctive characteristics shared by MRSA and CA-MRSA, respectively (Watkins et al., 2012).
MOLECULAR INSIGHTS OF ANTIBIOTIC RESISTANCE:
Because of its well-known ability to worsen the management of infectious diseases, surgery, and organ transplantation, antimicrobial resistance (AMR) in clinically relevant bacterial species has piqued the interest of researchers, clinicians, legislators, and executives from the pharmaceutical industry (Teillant et al., 2015). In addition, because antibiotics do not target particular pathogens, they may affect commensal microbiota that lives in the same environment as the treated organisms (Keeney et al., 2014). Because of the selective advantage they provide, AMR-producing genes are present in both pathogens and environmental and human commensal bacteria (Brinkac et al., 2017). Several in-silico metagenomics studies have identified hundreds of AMR genes in the genomes of commensals, symbionts, and opportunistic pathogens that inhabit the human gastrointestinal tract (Ghosh et al., 2013). Many antimicrobial drugs may have their efficacy reduced by AMR gene pools in the gut microbiota. This is because these pools might change the chemical composition of the medications and block them from reaching important microbial biological processes (Alekshun and Levy, 2007). Transduction and conjugation rely on the physiology of the donor and recipient bacteria, in contrast to spontaneous transformation, which relies only on the recipient cells' physical state (Forsberg et al., 2012). Transposition is the primary mechanism by which most naturally competent bacteria in the intestines acquire genetic material from other, closely related or unrelated, microbial species (Wellington et al., 2013). Bacteria may get resources or fitness traits via horizontal DNA acquisition, giving them an advantage over other species in the same ecosystem when conditions are bad (Finkel and Kolter, 2001).
Therapeutic strategies to combat antibiotic resistance:
Even if there is clinical evidence that treatments based on small-molecule monotherapy are ineffective in situations where resistance develops, the evolutionary race has always taken infection control seriously. Future research should primarily concentrate on identifying and discovering novel antibiotics, as shown by the examination of several candidates for infection control throughout development. Antibiotics, creative combination treatments, and drug delivery systems are developed at a slower pace than biotherapeutics, novel small molecules, and new medications, which are usually extensions of existing pharmaceutical classes (Brooks and Brooks, 2014). In a world where drug resistance has outpaced medicine development, systemic monotherapy treatments are inadequate. Thus, rather than fighting bacterial evolution, it may be more useful to use control strategies that seek out resistance mechanisms that are inspired by genetics. To prevent resistant bacteria from sensing their quorum, certain organisms, including red sea algae, use furanones (Pena-Miller et al., 2013). Biologics have the potential to combat multidrug resistance, and while their application in the treatment of bacterial infections is still in its early stages, this promise cannot be ignored. A more logistically focused search for a potential medicine candidate may be inspired by biology, but tiny molecules will always be essential in infection treatment (Baveja et al., 2004). Drug development tools for biologics should not be as extreme as those for small molecules in order to overcome biologics' drawbacks, such as limited controlled delivery options, incomplete in-vitro stability, inadequate high-throughput, complex screening tools, ineffective pharmacokinetics, and relatively unknown pharmacodynamics. Coincidentally, there is a great deal of promise at the intersection of biologics, molecules with a biological basis, and drug delivery systems (Choi et al., 2009). Targeting antibiotic resistance, eliminating susceptible bacteria, and protecting the human microbiome may therefore need a paradigm change comparable to that of complex viruses and cancer. Combining conventional antibiotics with innovative adjuvants and feasible limited delivery techniques allows for the development of such a strategy. It is possible that improved bioinformatics might be extremely useful in identifying new targets and achieving effective combination delivery, rather than relying on a number of expensive development models that often fail in clinical trials (Weber, 2014). Because it impacts both humans and other animals, combating antibiotic resistance requires a multifaceted approach. Despite the anticipated resolutions and the lack of suggested answers, virtually little has been done to put them into action thus far (Littmann and Viens, 2015). Antibiotics are now standard use in healthcare, and any disorganized effort would have devastating monetary, social, and medical consequences. As antibiotic resistance continues to rise, it seems that antibiotic efficacy is declining. Epidemiological trends in multidrug-resistant bacteria should be considered while developing treatment regimens for infection control. Theoretically, keeping the topic in the spotlight should encourage researchers to find ways to manage bacterial infections that are effective in preserving the host microbiome, identifying and preventing antibiotic resistance, and developing new antibiotics. Intelligent and local delivery tools, such liposomes, directed nanoparticles, and infection-responsive polymer-controlled distribution, as well as novel combination strategies, might be advanced with the use of leading methodologies that fulfill this need (Kumar et al., 2018) (Fig. 2).
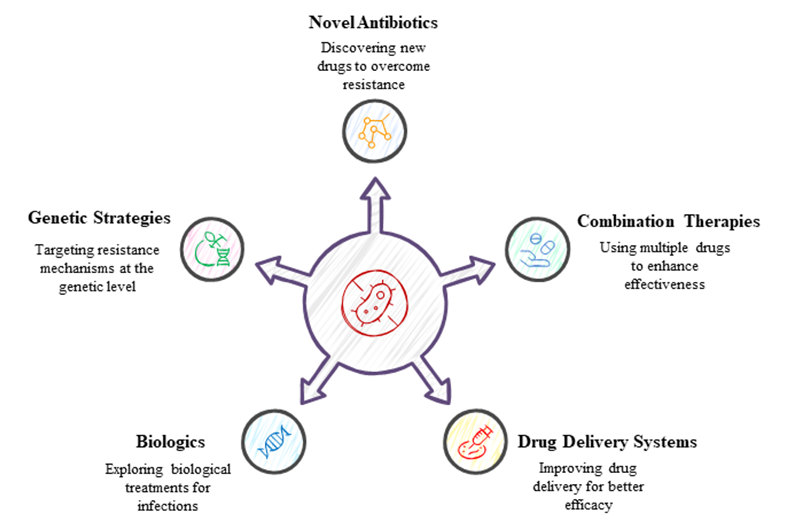
Figure 2: Different strategies to combat antibiotic resistance.
Consequences of the ABR crisis to human society:
In 2013, the "post-antibiotic" era was proclaimed by the US Centers for Disease Control and Prevention (Kumar et al., 2018). The AMR pandemic is worsening, according to WHO in May 2014. People will have to resort to less effective, but more traditional, means of infection management more often as a result of the rise of AMR in contemporary culture. Debridement, disinfection, amputation, isolation, and other invasive treatments will become more common, and treatment times will lengthen, as infections become less effectively treated. Not only non-lethal illnesses impact more people more often and take longer to cure, but the prospect of catastrophic global pandemics like the Spanish Flu of 1918 is not out of the question either. The increasing frequency of diseases that kill or seriously impair people will greatly affect human society (Kumar et al., 2018). Disease has always been a part of human history, but without efficient antibiotics, it threatens to wreak havoc on an already massive and ever-expanding human population. An increase in the prevalence of chronic, fatal infectious diseases would have a major effect on the economy because it will force more productive individuals to miss work for longer periods of time (Moore?Gillon, 2001). The increasing number of sick will also place a burden on healthcare systems generally and on the communities and families of those who are sick. The loss of skilled workers and the increased demand on healthcare systems will have a domino effect that reduces national production and impacts society and culture. After accounting for the cumulative impacts of increasing morbidity, the potential ramifications of incurable, fatal, viral pandemics would be comparable to, if not greater than, those of the 1918 global Spanish flu epidemic, which killed at least 50 million people (Trilla et al., 2008) (Fig. 3).
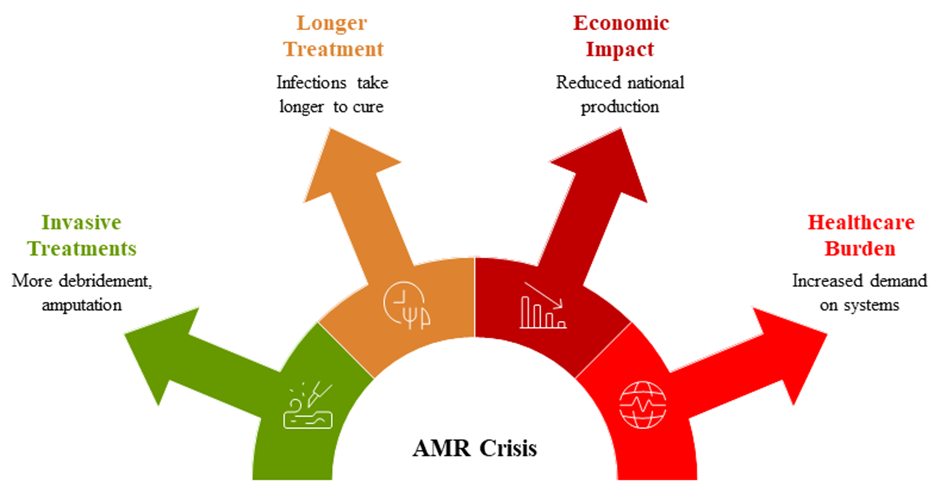
Figure 3: Impacts of antimicrobial resistance (AMR) crisis on human society.
Antibiotic resistance:
One of the most pressing issues in clinical practice right now is the rise of antibiotic resistance. The persistently alarmingly high levels of antibiotic resistance that many major infections, most notably S. aureus, exhibit have been recognized for some time (Witte, 1999). In addition, bacteria that form biofilms on prosthetic surfaces are often very resistant to antimicrobials and may withstand harsh chemotherapy, even when there are no known antibiotic resistance determinants. When this is considered, the need of testing for antibiotic-resistant bacteria in orthopedic therapeutic settings becomes clear, and not just when deciding on a course of therapy for the patient (Gold et al., 1996). These days, S. aureus and S. epidermidis are resistant to most beta-lactam medicines, especially penicillin. While a lower but significant fraction of bacteria exhibit signs of methicillin/oxacillin resistance, over 40% of germs in our orthopedic infection isolates associated with biomaterials are no longer susceptible to penicillin drugs like cephalosporins. There is less of an emphasis on methicillin/oxacillin resistance due to the fact that all b-lactam antibiotics are effective against methicillin-resistant bacteria. In addition, methicillin/oxacillin resistant staphylococci often exhibit multiresistance to several medicines from other antibiotic families, such as sulfonamides, aminoglycosides, macrolides, lincosamides, tetracyclines, and trimethoprim (Struelens and Denis, 2000). This means that, with the exception of vancomycin, all known types of antibiotics are susceptible to these two common staphylococci species. Despite the fact that our clinical context has never observed staphylococcal orthopedic clinical isolates resistant to vancomycin, the lack of adequate medical management to control the infection would be devastating if it did occur (König et al., 2001). In addition, our epidemiological data from systematically monitoring antibiotic resistance in all clinical isolates has recently shed light on several CoNS species that were previously ignored in numerous investigations. These species are typically rare among nosocomial infections, particularly those without implant materials, and they appear to have lower virulence and pathogenicity. Up until recently, most bacteria found in the central nervous system (CNS) were thought to be harmless, opportunistic pathogens. Consequently, they were sometimes grouped together under false names (Mah and O'Toole, 2001). Probably related to the different ecological niches that each species occupies, more data on the many specifics and antibiotic resistance features of each species is now accessible owing to rapid and accurate identification methods (Schwank et al., 1998). Since the prevalence of antibiotic resistance in S. haemolyticus exceeds that in the better famous S. aureus and S. epidermidis, it seems that this species is the greater cause for worry (Baquero, 1997).
The emergence of antibiotic resistance:
It is possible that an organism does not express its chromosomal determinants for drug resistance until it comes into touch with the drug or comparable substances (Sanders et al., 1988). Novel antibiotics or conjugative plasmids are two examples of favorable conditions that could lead to the rapid emergence of resistance (Chow et al., 1991). It is possible that the antimicrobial substance against which resistance is aimed may be the one that triggers its development. Sometimes, another antibiotic can cause the understudied antibiotic to become less effective by inducing or derepressing an enzyme or other factor that increases resistance to it (Sanders and Wiedemann, 1988). In every given population, there may be some strains that are resistant to the antibiotic being studied. When the bulk of the organisms in the population are susceptible, a stimulus that inhibits or kills them allows the resistant subset of strains to flourish (Party, 1989). While the antibiotic resistance in a given subpopulation is the most common determinant in selection, other, related drugs may sometimes play a significant role as well. Organisms may gain a selective advantage from non-drug variables as well, for example, those that increase the activity of glucuronidation or acetylation (McGowan, 1983). A patient can get an infection from a number of different sources, including other patients, healthcare providers (e.g., ward staff), commercial products (e.g., antiseptics) that are contaminated, inanimate objects, or even just the transfer of genetic material from one organism to another. As with the above described sharp and less abrupt changes in resistance, this sixth transmission pathway may be seen as a "common denominator". A study out of a medical center in Indianapolis found that endemic bacterial cross-transmission in an intensive care unit environment is very unusual, and that the few instances of dissemination that did occur included diverse species (Chetchotisakd et al., 1994).
Drivers of antibiotic resistance transmission:
At this time, understanding the many causes of antibiotic resistance is crucial to finding a solution to this global problem. Natural selection has kept germs resistant to antibiotics at bay until recently. In today's world, antibiotic resistance is being actively promoted by a variety of medications used in agriculture, livestock, healthcare systems, and the environment. Medication quality, sanitation and water systems, infection control protocols, medicines and diagnostics, and migratory or transit quarantines are other important variables that greatly contribute to the development of antibiotic resistance. Mutations in several genes located on the chromosome of the bacterium and the exchange of genetic material across organisms are the main causes of antibiotic resistance (Holmes et al., 2016). Plasmid transmission is the most important event that might transfer genes that make cells resistant to antibiotics to the host cell. The distribution of antibiotic-resistant components is one-way antibiotics can influence this process; another is that they might exert a selection pressure that prevents antibiotic resistance from developing (Munita and Arias, 2016). Since the transmission patterns of resistance have been shown, our understanding of how these illnesses transmit from one person to another has expanded (Laxminarayan et al., 2013). In communities, the feco-oral route is the most important vector for the spread of resistant infections, especially those in the Enterobacteriaceae family. This usually happens when there isn't enough cleanliness. The mechanisms of resistance transmission between people may be better understood by looking at community-acquired (CA)-MRSA. This kind of MRSA is often disseminated due to long hospital stays or dirty hospital surroundings. Furthermore, sexual contact may transmit N. gonorrhoeae that are resistant to treatment (Wellington et al., 2013). Another important factor, "one health", greatly affects the dynamics of antibiotic resistance transmission. Considerable infections, such as Salmonella and Campylobacter species, may be transmitted to humans via animal products due to the excessive use of antimicrobial growth promoters in livestock. Also, comparable resistance mechanisms have been seen in bacteria and viruses isolated from animals and humans. Mobile genetic elements (MGEs) and resistant bacteria may transfer from animals to humans in a number of different ways (Mølbak, 2004). How environmental factors amplify antibiotic resistance is another cause for concern. The agricultural business uses metals as microbicides, and these metals might be a major factor in the development of resistance. Moreover, it is well-known that sewage systems, pharmaceutical industry pollution, and waste management practices all contribute to the transmission of illnesses from humans to other organisms. Both pre- and post-treatment analyses of sewage systems have shown the presence of many resistant bacteria (Kristiansson et al., 2011) (Fig. 4).
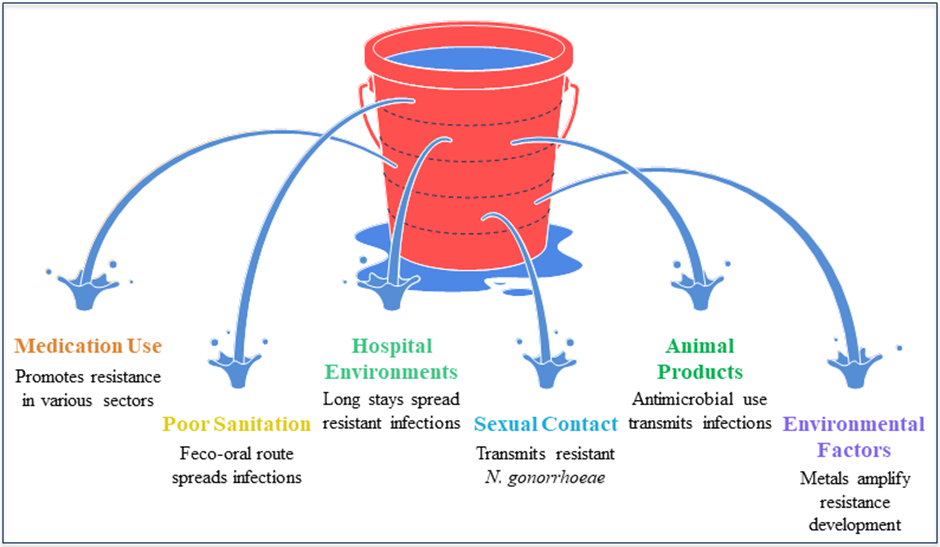
Figure 4: Different drivers of antibiotic resistance transmission.
Origins of antibiotic resistance:
No two bacterial species or groups will always have the same level of susceptibility to or resistance to the same antibiotic. Even among groups of bacteria that seem to be somewhat similar, resistance levels could vary greatly. The minimum inhibitory concentration (MIC) is the concentration of a drug at which further bacterial growth is inhibited, and it is the standard unit of measurement for determining sensitivity and resistance (Coculescu, 2009). In reality, antibiotic susceptibility is defined as the range of average MICs for a given antibiotic within the same bacterial species. If a species' average MIC is in the range of values that indicate inherent resistance to a treatment, we say that species is resistant. The level of resistance that bacteria acquire varies not just from one species to another, but also from one level of similarity to another (Martinez, 2014). Bacteria display two forms of inherent resistance: intrinsic resistance, which is constant across species, and induced resistance, which is maximally developed in response to antibiotic treatment. Intrinsic resistance is a trait that all bacteria have, it has nothing to do with horizontal gene transfer, and it is unaffected by previous exposure to antibiotics (Cox and Wright, 2013). The most common bacterial mechanisms implicated in intrinsic resistance are reduced permeability of the outer membrane (particularly the lipopolysaccharide, or LPS, in gram negative bacteria) and the inherent functioning of efflux pumps. In addition, one common way of induced resistance is by multidrug-efflux pumps (Fajardo et al., 2008). Bacteria may acquire resistance-inducing genetic material by any of the three main mechanisms of horizontal gene transfer (HGT): transposition, conjugation, and transformation. The bacteria's own chromosomal DNA might also change. The duration of the purchase might be either brief or lengthy. Although bacteriophage-borne transmission is rare, plasmid-mediated transfer of resistance genes is the most prevalent way of obtaining foreign genetic material. Acinetobacter species and other bacteria with intrinsic competence may be able to directly absorb genetic material from their environment. Bacterial genetic changes (such as deletions and substitutions) may occur in response to environmental challenges, such as chemicals, ultraviolet radiation, starvation, and insertion sequences, which can rearrange genetic material. Bacterial mutations often result in cell damage; on average, there is one mutation for every 106 to 109 cell divisions (Davies and Davies, 2010). Also, there are a lot of modifications that cause antibiotic resistance, and the organism has to pay for them. When Staphylococcus aureus bacteria gain resistance to methicillin, for example, their growth rate drops significantly (Reygaert, 2008). Antimicrobial resistance is a big enigma, and one of the biggest mysteries is that these drugs make resistance worse. Using sub-inhibitory concentrations of antimicrobials, even at low ones, can have several negative effects. For example, it can promote the selection of highly resistant bacteria in future generations, increase the rate of mutation, make bacteria more resistant to other antimicrobials, and even facilitate the migration of mobile genetic elements (Blázquez et al., 2012).
The ABR mechanism:
A large amount of literature on the molecular bases of antibiotic resistance has examined the biochemistry and genetics of several bacterial cell activities (Tsui et al., 2004). Research on antibiotic action and resistance has actually improved our knowledge of cellular structure and function. The vast majority of resistance mechanisms include several vectors for gene transmission. There is a great deal of evidence that resistance mechanisms are present in many different types of microbes, including both commensals and pathogens. Some forms of resistance stand out as particularly egregious examples of the difficulties associated with maintaining antibiotic effectiveness in the face of bacterial genetic and biochemical adaptation (Marshall et al., 2009). The genes responsible for generating -lactamases are probably the most common; catalysts with an ever-expanding resistance range have been created from random mutations in these genes (Gniadkowski, 2009). The fact that many closely similar enzyme families have evolved from the prototypical plasmid-encoded lactamase (TEM) is evidence of its adaptability. The presence of unique -lactamases with changed substrate ranges in the environment is likely due to the fact that these genes are ancient and have been found in isolated, desolate locations. To provide an extra example, environmental Kluyvera strains in the 1990s were the first to develop a new extended-spectrum \-lactamase (CTX-M), the first enzyme to hydrolyze expanded-spectrum cephalosporins to a clinically actionable level (Livermore et al., 2007). The CTX-M genes are a problem all around the globe, and the variations of these genes—which have up to 100 different amino acid changes—are spreading rapidly. It is quite unlikely that efficient HGT and rapid mutational radiation would be able to control such gene outbreaks (Hawkey and Jones, 2009). Methicillin resistance prompted the development of macrolide antibiotics, which are widely used to treat Gram-positive infections. The antibiotic erythromycin and its variations are examples of this class of drugs. Unsurprisingly, there has been a substantial increase in the distribution of strains that are resistant to different approaches recently (Roberts, 2008). The macrolides and similar antibiotics bind to different spots inside the peptide escape tunnel of the 50S ribosomal subunit, which is how they exert their action. Alterations to the tunnel's RNA or protein components can cause resistance. A specific rRNA mutation that confers resistance to all antibiotics acting at this ribosome site has just been found, and this mutation is rapidly expanding (Long et al., 2006). Intrinsic resistance occurs when a bacterium's genome contains genes that might lead to a resistance phenotype, such proto- or quasi-resistance. Various genera, species, strains, etc., exhibit phenotypes that vary in their responsiveness to antibiotics. Many potential or intrinsic gene functions in bacteria have been revealed after the year 2000 because of advancements in genome wide mutagenesis technologies and rapid bacterial genome sequencing. Some of these activities may cause resistance phenotypes in medical contexts. For example, antibiotic resistance, especially to sulfonamides, may develop by gene amplification, a common genetic mechanism (Kashmiriand Hotchkiss, 1975). Hints on possible future advancements are provided by these studies. By conducting phenotypic studies on partial or "complete" gene knockout libraries, specific mutants that cause antibiotic-induced hypersensitivity responses may be identified via saturation mutagenesis of bacterial genomes. The overexpression of the specific wild-type gene is believed to provide a resistance phenotype. These prognostic studies have forecasted the rise of novel resistance classes in a wide variety of species. The first study of this sort used a library of mutant strains isolated from Acinetobacter baylyi (Gomez and Neyfakh, 2006). Using a mutant approach, Yassin and Mankin found possible sites to insert inhibitors of ribosomal activity. Research into rRNA has uncovered a plethora of potential new targets for small-molecule translation inhibitors by identifying RNA segments. Innovative research like this demonstrates that, despite seemingly contradictory expectations, the antibiotic development sector still has a wealth of untapped medicinal potential. Antibiotic lifetime extension might be achieved by the use of reliable resistance prediction and prompt response (Martínez et al., 2007).
The ABR, a growing public health threat:
An increasing number of global health organizations are starting to recognize antimicrobial resistance (AMR) as a global public health crisis, a problem with our present healthcare system, and a threat to modern medicine and the control of infectious diseases. A growing body of evidence suggests that "selective pressure" due to antibiotic overuse in humans and animals is a major contributor to the worldwide AMR pandemic (WHO, 2001). There is a correlation between infections caused by antibiotic-resistant strains and a decline in quality of life, an increase in the likelihood of recurrence, chronicity, infections spread to other parts of the body, and the development of resistance organisms via opportunistic infections (Helms et al., 2004). Evidence of this issue is provided by the increasing resistance of human pathogens, which includes pathogens like Salmonella, Campylobacter, and vancomycin-resistant enterococci, some of which are fatal. Additionally, there is a high rate of treatment failures, complications, worsening of pathological conditions, and death (IDSA, 2011). Antimicrobial resistance is estimated by the European Centre for Disease Control and Prevention (ECDC) to be responsible for 25,000 yearly deaths and 2.5 million extra hospital days in the EU, Iceland, and Norway, according to 2009 estimates. Similarly, AMR was responsible for 100,000 deaths in the United States and 80,000 deaths in China over the same time frame. Regarding nosocomial infections caused by bacteria with resistance, Margaret Chan, the head of the World Health Organization, said that the "post-antibiotic era means, in effect, an end to modern medicine as we know it" (Chan, 2012). Recurring causes of mortality include common illnesses such as strep throat or a skinned knee in children. The proliferation of AMR bacteria is seen as a significant danger to public health, with consequences that might be compared to those of climate change and other social and environmental dangers. In its 2014 Global threats research, the World Economic Forum evaluated fifty external global threats according to their probability, impact, and linkages; the risks were then rated according to their potential implications on the economy, environment, geopolitics, society, and technology (Spellberg, 2008). We compared the impact and probability of AMR to those of terrorism and global warming.
Economic cost of ABR:
Instability and increased costs for health systems could be consequences of antimicrobial resistance. A longer recovery duration, a larger prevalence of septicemic infections, and a higher mortality rate are all experienced by individuals who get antimicrobial resistance nosocomial infections (mostly bloodstream infections) or become ill from consuming food contaminated with these germs (Angulo et al., 2004). Health care costs are higher in this case due to the lengthier hospital stays and more costly drugs that are used. Concerns about toxicity, adverse drug reactions (ADRs), and collateral events are all on the rise with the introduction of new medications. Health care costs, both direct and indirect, were estimated to be roughly €1.5 billion per year, with lost productivity amounting to €600 million for the European medical health business. The AMR has a larger clinical impact than HIV infection and is estimated to cost the US economy more than $55 billion a year in healthcare costs. Healthcare costs are expected to be disproportionately high due to drug-resistant germs, according to economists, who warn that reporting system deficiencies impact mortality and morbidity data (Smith and Coast, 2013). The situation has worsened because to the economic and financial crisis that swept over Europe in 2008 and beyond, which may lead to a decrease in funding for public healthcare and investment in novel compound development by pharmaceutical companies. These socioeconomic determinants have an inseparable effect on the epidemiology of antimicrobial resistance nosocomial infections. It is worth noting that low-income countries are believed to pay a disproportionate share of healthcare expenditures because of the negative effects of inadequate infrastructure, health care, and economic resources on medication availability and proper utilization (Sosa et al., 2010) (Fig. 5).
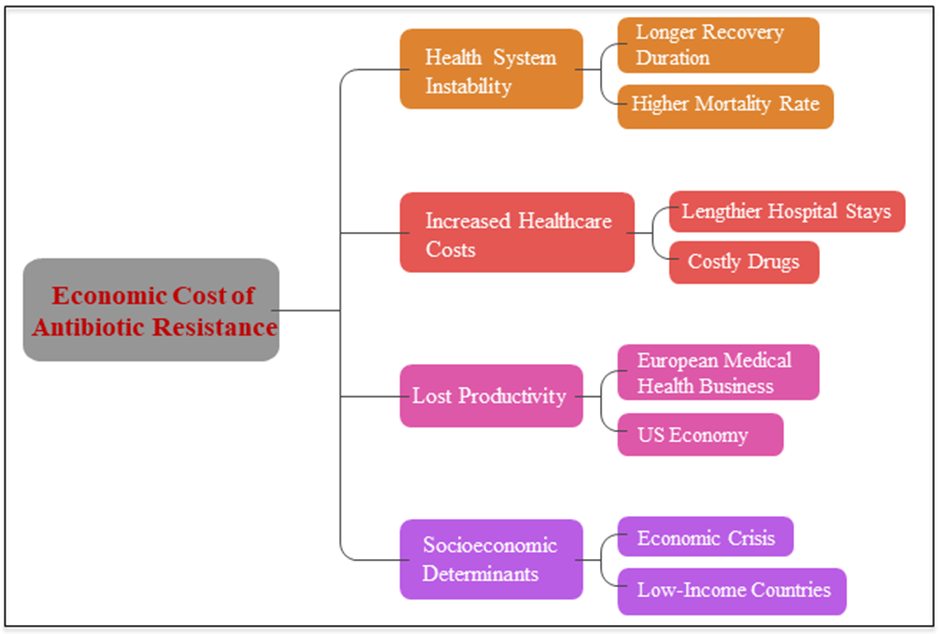
Figure 5: A number of economic impacts of antibiotic resistance.
The ABR in food chain:
Several resistance illnesses in farm animals and other foods generated from animals are undeniable (Oliver et al., 2009). Because of the complexity of the many routes of transmission within and between species, as well as the frequent transfer of resistance genes among host bacteria, it has been difficult to formulate a definitive answer to the question of whether antibiotic use in food-producing animals has contributed to the emergence, spread, and persistence of AMR in humans. This is true even though there is abundant evidence that animals and the environment are major contributors to bacteria with resistance to antibiotics, and that people are at danger as a result (Wielinga and Schlundt, 2012). Multiple lines of evidence have recently emerged connecting antibiotic usage in cattle (both therapeutic and nontherapeutic) to the development of antibiotic resistance in humans. These links have been shown in studies using ecological, historical, microbiological, and controlled subject data (including disease outbreaks) (Chapin et al., 2010). More powerful and up-to-date epidemiological techniques are molecular genetic approaches, which may reveal the presence of the same gene (or plasmid) in strains from various species, whether they are animals or humans. Molecular typing and phenotypic data are now the best study tools for determining whether resistant isolates found in food or animals are same to those employed in human healthcare. Mathematical models may be used to quantify the contribution of each main animal food source to human salmonellosis caused by bacteria that are resistant to antibiotics (Hald et al., 2007). From a quantitative standpoint, food is the most important vector for the transmission of antibiotic-resistant genes and bacteria from farms to consumers. Eating foods derived from animals puts people at risk of contracting bacteria that are resistant to antibiotics, according to a plethora of research. Alexander et al. (2010) found drug-resistant Escherichia coli on the surfaces of beef carcasses during evisceration, after 24 hours in the refrigerator, and in ground beef that had been stored for one to eight days. Campylobacter spp. resistant to ciprofloxacin was found in chicken products consumed by the general public in the United States. The Italian town of Normanno found MRAS in cow's milk (Normanno et al., 2007). One study found that 20% of 200 samples of ground beef sold in stores contained Salmonella. A whopping 84% of the Salmonella strains tested showed resistance to at least one antibiotic, with 50% showing resistance to three or more. Concerningly, the drug ceftriaxone is suggested for the treatment of severe Salmonella infections in children, but 16% of the isolates were resistant to it (White et al., 2001).
Antibiotic resistance in the environment:
New evidence suggests that many different types of bacteria may exist in nature, some of which have resistance genes and may cause illness under the correct circumstances. The proportion of soil bacteria harboring antibiotic resistance genes has increased significantly since 1940, according to a research conducted in the Netherlands (Knapp et al., 2010). Approximately 27% of the samples collected from New York City subway stations included live, antibiotic-resistant bacteria, and one station even yielded a multi-drug-resistant culture (Afshinnekoo et al., 2015). This finding was made by researchers in the United States who recently researched microorganisms. It's possible for drug-resistant bacteria to infect humans and other animals via a number of environmental vectors, such as food crops cultivated on soil that has been treated with antibiotic-containing manure. Community exposure to crop fields where swine manure was used as fertilizer is a significant risk factor for both health care-associated and community-associated MRSA strains, according to a case-control study investigating the relationship between environment-resistant pathogens and human health that was carried out in Pennsylvania from 2005 to 2010. The study included over 400,000 primary care patients (Casey et al., 2013). The environment around livestock farming operations has been discovered to contain antibiotic-resistant bacteria that originate from animals. The bulk of the streams with antimicrobial residues were located downstream from animal agriculture companies (Kolpin et al., 2002), and 48% of the 139 streams analyzed nationwide were part of this study. Researchers found that cattle grazing close to water sources may pollute aquatic life with antibiotic-resistant diseases like E. coli. One thousand samples of salads, fruits, roots, and bulbous veggies were examined by researchers from Germany. They found that common plant bacteria like Pseudomonas and Enterococcus contained resistance genes that might be transmitted to bacteria in the digestive tract (Schwaiger et al., 2011).
Use and overuse of antibiotics in populations:
Using antibiotics on animals for food makes an already serious problem much worse, and the problem seems to be worse in developing countries. In both the 2003 publication and the 2009 study, the World Health Organization (WHO) made the same claim. Stressed how the misuse and overuse of antibiotics in livestock production negatively impacts public health. There are "major gaps" in data sharing and monitoring about the impacts of antibiotic-resistant bacteria in food on both people and animals, as pointed out in the 2009 Global Report on Antimicrobial monitoring. One gap that has to be addressed is the amount of antibiotics used each year by animals that are grown for food throughout the globe (Thauvin?Eliopoulos and Eliopoulos, 2003). Antibiotic use in animal husbandry is rampant, according to many countries. A recent study found that out of China's 210,000 tons of antibiotic production, almost 50,000 tons are applied to animals raised for food. Over 80% of all antibiotics were used on animals grown for food, which is quite discouraging according to statistics from the United States (Dalhoff and Schmitz, 2003). Since most antibiotics are not broken down in animals raised for food, releasing 75–90% of them into the environment is a major consequence of antibiotic overuse. These drugs have a lengthy half-life in the environment, which means that bacteria that come into contact with them have plenty of opportunity to evolve resistance (Keddy et al., 2010). By mapping it, we may get a baseline estimate of the global relevance of antibiotic ingestion in cattle. A recent research published in the Proceedings of the National Academy of Science (PNAS) provides the first worldwide map of antibiotic usage in cattle internationally (228 nations) (Van Boeckel et al., 2015). The study's projections show a cumulative rise of 67% from 2010 to 2030, which might put human antibacterial effectiveness at risk. Global estimates for the proportion of antibiotics used on animals place China and the United States at the top, based on data between 2010 and 2030. A rise of 99% in antibiotic usage is projected in the BRICS countries: South Africa, Brazil, Russia, India, and China (Fig. 6). While both the Chinese and Indian governments have made strides in curbing antibiotic misuse by the human population, they have lagged behind when it comes to regulating antibiotic use in animal feed (Bax et al., 2001).
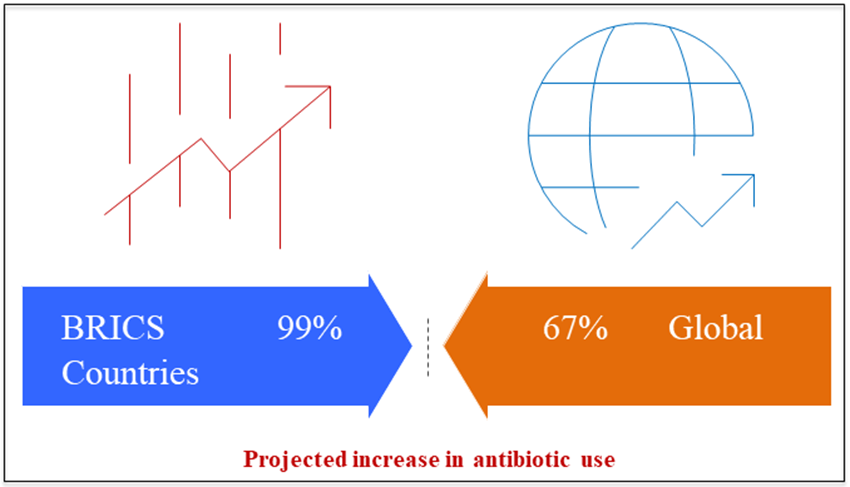
Figure 6: Antibiotic use in animals (2010-2030) by countries of world wide.
Reduce antibiotic resistance development:
Antibiotic regulation initiatives have persisted since the simultaneous growth of resistance bacteria is the most serious adverse outcome of antibiotic use. A good example of this is the early antibiotic erythromycin, which was withdrawn off the market entirely within a year of the discovery that 70% of S. aureus isolates had become resistant to it. Boston City Hospital began using erythromycin in the early 1950s to treat Staphylococcus aureus instead of penicillin. Following its discovery with chlortetracycline and chloramphenicol, the same phenomenon was seen with other antibiotics (Stelling et al., 2005). The emergence of antibiotic resistance is a clear and present danger. How do we put a halt to this, or at least make it go more slowly? Over the years, several different techniques have been proposed by knowledgeable experts and all of the major international health agencies, such as the CDC and the WHO. Antibiotics should not be used to treat viral infections or colds; there should be stringent regulations on their use by humans, including the requirement for accurate prescriptions; a ban on the delivery of antibiotics without a prescription to reduce their unnecessary use; and restrictions on their therapeutic use in animal husbandry and agriculture (Connell et al., 2003). It is noteworthy to note that the 1969 Swann recommendations were the pioneers in calling for the prohibition of non-therapeutic use in livestock and farming. Despite its reasonableness, this plan caused division and is still not enforced in many nations. Several of the antibiotics that have been approved for human use are given to animals under different names for different purposes, according to a report by the Alliance for the Prudent Use of Antibiotics, which suggests that deceit played a role in this failure (Aminov et al., 2001). It is clear that implementing limitations on antibiotic consumption on a global scale is problematic, despite the fact that resistance levels have been effectively lowered in the Netherlands and Scandinavia. Is it possible that resistance will go away if everyone follows the suggested restraint protocols? In all likelihood, no. The most recent (among many) papers on the topic is Antibiotic Resistance: An Ecological Perspective on an Old Problem (Roberts, 2005). Antibiotic resistance is most often caused by the ability of most environmental bacteria and their pathogenic counterparts to pump antibiotics out of cells. For the purpose of developing individualized or combination drugs, it is appealing to find compounds that block the outflow of active inhibitors from the cell (Chopra and Roberts, 2001).
RESISTANCE SURVEILLANCE STUDIES:
Fluoroquinolone:
Infections in both inpatient and outpatient settings are often treated with fluoroquinolones nowadays. Despite the high number of prescriptions for quinolones throughout the first 20 years of its availability, drug resistance during treatment was uncommon apart from Staphylococcus aureus, particularly methicillin-resistant Staphylococcus aureus (MRSA) and Pseudomonas aeruginosa. Two species rapidly evolved resistance to fluoroquinolones, mostly via the spread of genetically similar strains among immunocompromised people and those residing in nursing homes (Bryan et al., 2004). However, quinolone resistance has been steadily increasing across the board for both Gram-positive and Gram-negative bacteria since the mid-1990s. Concerningly, resistance rates continue to climb (Altenbuchner et al., 1983). Due to the prevalence of serious side effects, the warning portions of the package inserts for norfloxacin (1986), ciprofloxacin (1987), levofloxacin (1996), and moxifloxacin (1999) have undergone several adjustments since their US approval. Fluoroquinolones are no longer used without permission according to the European Medicines Agency (EMEA). Since July 4, 2008, there have been restrictions on the usage of oral formulations of norfloxacin. There is a lack of evidence showing that oral therapy with drugs containing norfloxacin is useful in complex pyelonephritis, which led to this conclusion. Nevertheless, the previously given clearances have remained unchanged despite changes in the susceptibility patterns of the disease-causing bacteria. Thus, this research aims to show that patients' care is affected and that certain current treatment plans need to be adjusted due to the persisting increase in fluoroquinolone resistance rates (De Baere and De Backer, 2007). Important trends in the spread of infections and the evolution of antibiotic resistance may be detected at sizes varying from the local to the global thanks to the data collected by surveillance studies. Antimicrobial sensitivity testing has been, and is still being, the standard practice. However, many studies that included monitoring people have been shown to have biases or errors that have not been corrected. Some examples of such restrictions include narrowing the emphasis to a certain region, collecting data from a subset of patients, or limiting the study's geographic reach to a given country, state, or even neighbourhood. Furthermore, user needs may not be well defined; for example, infection control experts and microbiologists may be interested in collecting data on resistance incidence or prevalence (Yang et al., 2002). In order to detect and monitor the spread of resistance, surveillance studies are essential. As a second line of defense, carbapenems, aminoglycosides, and piperacillin/tazobactam are the main subjects of hospital-based surveillance studies. Contrarily, fluoroquinolones are more often prescribed by primary care physicians than by hospitals. Because of the differences in fluoroquinolone prescription patterns between primary care physicians and specialists, as well as between patient demographics and bacterial species, the second goal of this study is to classify the collected data accordingly. Since only data from monitoring programs will be taken into account, the question of how fluoroquinolone resistance has emerged in species like Clostridium difficile, Salmonella spp., and others will remain unanswered (Valsamatzi-Panagiotou et al., 2020).
Tetracyclines:
Chlortetracycline, minocycline, doxycycline, and tetracycline are the four main antibiotics that make up the tetracycline family. Atypical compounds include anhydrotetracycline and 6-thiatracycline (Yao et al., 2019). Conventional tetracycline blocks the attachment of aminoacyl-transfer RNA (tRNA) to the 30S component of the ribosome, which inhibits protein synthesis (Higgins et al., 2004). Infections in humans, animals, and plants may all be treated with these bacteriostatic drugs. Medications used in whole or in part to prevent bioterrorism-related infections such as Francisella tularensis, Yersinia pestis, and Bacillus anthracis are additional indications (Runyon et al., 1987). By combining DNA hybridization with restriction enzyme analysis and tetracycline resistance gene expression analysis, Mendez was able to classify genetic variables linked with resistance in Enterobacteriaceae in 1980. There are many mechanisms via which tetracycline resistance develops, including as ribosome protection, efflux pumps, altered enzymes, and changes in drug targets. These findings point to the involvement of additional plasmid genes in this resistance mechanism, including tet and otr. The development of resistance to oxytetracycline is influenced by three different genes. There has been an inhibition of minocycline and doxycycline activity by the ribosomal proteins (Runyon, 1988). The first bacterium known to be resistant to tetracycline was Shigella dysenteriae in 1953. This resistance gradually expanded to other species as time passed. Typically, at least one of the 36 known tet genes is responsible for tetracycline resistance. Efflux pumps, ribosome protection, and drug target alteration are the three main ways these genes act to resist drugs. Both Gram-positive and Gram-negative bacteria often have ribosome protection mechanisms and the efflux pump (Dupeyron et al., 1988). All of the known tet genes are involved in the ribosome protection and efflux pump systems, however only the tetX gene is involved in the enzymatic modification mechanism. Extensive research into the process of tetracycline resistance via the transposon Tn10 has shown that a structural gene produces the 36,000 Da resistance protein, and a regulatory gene encodes a repressor protein with a molecular weight of 250,000 Da. The tetA gene codes for resistance and the tetR gene codes for repressor proteins; both are located on the Tn1721 transposon. The activation of these genes is enhanced by tetracycline (Navasa et al., 1996).
Amoxicillin:
Bacteria develop resistance to antibiotics when they continue to multiply even when exposed to these drugs. Numerous strategies that are clinically meaningful allow bacteria to maintain their resistance. Antibiotic resistance may evolve in microbes in a variety of ways, including genetic changes and non-genetic mechanisms (Felisart et al., 1985). Antibiotic detoxification, internal antibiotic accumulation, drug target substitution, and protection are some of the mechanisms by which bacteria acquire resistance. The concentration of amoxicillin in the aquatic system is 6.94 μg/L. The major concern is that extended use of amoxicillin might cause the emergence of genes that are resistant to the drug (WHO, 2004). Essential components of both Gram-negative and Gram-positive bacteria are peptidoglycan polymers; amoxicillin, a broad-spectrum antibiotic, inhibits their cross-linking. There are many mechanisms by which the bacteria overcome the amoxicillin stress. The principal process in Gram-negative bacteria is the creation of Beta-lactamase. The Metallo-Beta-lactamase enzyme is secreted by gram-negative bacteria and has a broad range of action. The beta-lactamase inhibitors sulbactam and clavulanic acid work synergistically to increase amoxicillin's efficacy, especially against bacteria that produce the enzyme. The spectrum of amoxicillin's effect is therefore expanded (Wu, 2012). Bacterial resistance to amoxicillin's beta-lactam ring develops when beta-lactamase inhibitors permanently bind to the active site of the enzyme. Beta-lactamase production, plasmid-mediated AmpC enzyme transfer, and the existence of ESBLs (extended-spectrum beta-lactamases) like blaTEM, blaSHV, blaCTX-M, and carbapenemases are the mechanisms by which beta-lactam antibiotic resistance develops. The carbapenem-resistant Stenotrophomonas maltophila has the metallo beta lactamases (MBL) L1 gene. As a result of alterations to the penicillin-binding proteins (PBPs) and a decline in the synthesis of outer membrane proteins, antibiotic penetration is reduced in gram-positive bacteria (Van Boeckel et al., 2015). One new protein, PBP 2a, is formed in part by the antibiotic binding sites found in the PBPs. Because it helps bacteria make their cell walls, this protein is essential for resistance development. Both the structure and metabolism of peptidoglycan are affected by the presence of femX in S. aureus. This causes the cell membrane to abnormally thicken, which eventually makes the bacteria resistant to amoxicillin. The development of antibiotic resistance in stressful situations is also influenced by many genes that have not yet been discovered. Ulcers and cancer of the stomach may be caused by Helicobacter pylori. Some antibiotics, such rifampin and amoxicillin, are no longer effective against H. pylori (Rosenblatt-Farrell, 2009). In order to become resistant to amoxicillin, H. pylori may decrease the absorption of beta-lactams and the capacity of the PBP1 protein to bind to the antibiotic. Medications that block the action of beta-lactamase include sulbactam and Augmentin, which is a combination of amoxicillin and clavulanic acid with a brand name. On the other hand, Augmentin-resistant bacteria may produce beta-lactamase, an enzyme that degrades Augmentin's clavulanic acid. Although it is a beta-lactamase inhibitor, Augmentin is more efficient against Staphylococci than sulbactam. Tazobactam, a sulbactam analogue, has inhibited beta-lactam activity just as well as clavulanate (Collignon and Voss, 2015).
Cephalosporin:
Patients with severe cirrhosis are at increased risk for developing spontaneous ascites infection (SAI), a serious consequence. It is the main cause of liver transplantation and also a risk factor for secondary complications such hepatorenal syndrome and variceal hemorrhage. Because it has shown a greater success rate in identifying bacteria in the ascitic fluid compared to standard approaches, the bed-side blood culture bottle approach is becoming more recommended by gastroenterologists (Finland, 1979). It may be possible to start effective therapies for SAI more quickly if we knew which bacteria were causing the infection and how susceptible they were to medications. The majority of infections are caused by Gram-negative aerobic bacteria that are members of the Enterobacteriaceae family or by non-enterococcal Streptococcus species. Gram-positive bacteria that are resistant to antibiotics are beginning to create serious health problems. The impact of antibiotic resistance on patient outcomes has been the subject of very few studies (DeVincent and Viola, 2006). Empirical antibiotic therapy with high activity against the most typically observed organisms in SAI should be initiated before the decisive results of ascitic fluid cultures are known. Permanently deadly adverse effects may occur as a consequence of therapy inadequacies, treatment failures, or treatment delays. It is important to regularly update the spectrum of harmful microorganisms in a population and their antibiotic susceptibility. Evidence suggests that some oral antibiotics are beneficial in treating spontaneous bacterial peritonitis (SBP), which raises the question of whether or not they are cost-effective (Acar et al., 2009). This prospective study aimed to compare the empirical efficacy of cefepime, a fourth-generation cephalosporin, with cefotaxime, the most commonly used antibiotic supported by well-designed studies, and to identify sensitivity to various oral and parenteral antibiotics. It also aimed to explain potential changes in the microbial spectrum of SBP in cirrhotic patients (Lomovskaya et al., 2007).
CONCLUSION:
Finding out how widespread the AMR epidemic is should be our top priority. Accurate and thorough data collection is crucial for AMR regulation. The present state of worry around this subject is caused by a lack of sufficient understanding. No one can say for sure what the future holds, but with no new medicines on the horizon, it seems like AMR management will be a major challenge. The best way to tackle this issue is to employ a combination of techniques. Continual, interesting education for pharmacists, physicians, and medical students is essential. The policy should include rules that make it mandatory to closely track how often antibiotics are used. The development of novel diagnostic and screening tools requires an international and interdisciplinary team effort. We must not ignore the ecological and environmental aspects of the issue; rather, our management strategy must include all aspects of "one health." Other methods might also work, especially in countries with lower incomes. It is clear that AMR is no longer an unimportant issue, given the amount of focus on it. A global code of conduct that combines all existing efforts to eradicate AMR may, in the long run, succeed where this approach has failed. Two antimicrobial options that might reduce the global effect of AMR are lytic bacteriophages and probiotics. The AMR may be mitigated by a combination of measures, including immunization, infection prevention, smart antibiotic use, screening, treatment, education, and awareness of the disease among the general public. National, regional, and worldwide efforts to control the "global resistome" may benefit from bio-surveillance, response, prevention, and monitoring of AMR and MDR infections. Implant materials are associated with the majority of post-operative infections in orthopedics and many other medical fields because they allow even low-virulence bacteria to easily establish themselves by weakening the human immune system. Reducing the likelihood of infections after surgery is crucial since the number of patients using implant materials is enormous and rising rapidly. An increasing number of initiatives are aiming to reduce the implant materials' inherent susceptibility to microbial colonization, in addition to limiting the use of systemic antibiotics during surgery and the contamination of the surrounding environment and personnel. Improvements in the biocompatibility of implant materials are just as important as this area of technological innovation, which is a key focus in orthopedics. Since research on anti-infective surfaces is still in its infancy, it is reasonable to assume that funding and effort will soon expand in this area.
DECLARATION OF COMPETING INTEREST:
The authors assert that they do not have any conflicts of interest.
ACKNOWLEDGMENTS:
The first author is grateful for the support from the Global College of Pharmaceutical Technology, Krishnanagar-741102, West Bengal, India and corresponding author [BS] is acknowledging the support of Karimpur Pannadevi College, Karimpur-741152, West Bengal, India.
REFERENCES


Bikram Sarkar, Bipul Sarkar, A Comprehensive Study on Approaches in the War Against Antibiotic Resistance, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 1, 1729-1756. https://doi.org/10.5281/zenodo.18280525
 10.5281/zenodo.18280525
10.5281/zenodo.18280525
