Department of Pharmacology, Swamy Vivekanandha College of Pharmacy, Elayampalayam Tiruchengode - 637205, Namakkal.
A systemic skeletal illness called osteoporosis is characterized by a decline in bone mass and a breakdown of bone microarchitecture, which makes bones more brittle and more prone to fractures. Osteoporosis is classed as primary or secondary based on its etiology. Researchers use hip fractures to measure the severity of osteoporosis. This disease is responsible for the majority of hip fractures. Phytoestrogens have been examined extensively for their ability to prevent and treat aging-related disorders, including menopausal symptoms, skin aging, cardiovascular, neurological, immunological, metabolic, and cancer. The phenylpropanoid pathway produces phytoestrogens, a significant category of plant chemicals. Enzymes convert these molecules into various chemical forms. Plant compounds with unique structures can interact with estrogen receptors, resulting in a diverse range of phytoestrogens. Phytoestrogens are categorized as is flavonoids, flavonoids, chalcones, flavanones, stilbenes, and lignans. The development of bone resorption caused by osteoclastogenesis, reduced mineralization of bone tissue and the formation of bones due to reduced function of osteoblasts resulting from apoptosis and age-dependent differentiation of osteoblast precursors into adipocytes, and inflammation, oxidative stress, cell death, and epigenetic factors were all identified in experimental and clinical studies aimed at examining the primary pathogenetic mechanisms of osteoporosis. This study investigates the mechanics, efficacy, and clinical potential of phytoestrogens in controlling postmenopausal osteoporosis, providing insights into their role as a natural alternative to traditional hormone replacement therapy.
Osteoporosis is a popular skeletal disease characterized by decreased bone strength owing to impaired bone structure and an increased risk of fracture. Osteoporosis is a major source of concern in the elderly, and the severity of osteoporotic fractures increases with age, making treatment more difficult. As a result, it impacts the physical, psycho-social, and financial aspects of life for humans. Osteoporosis and osteoporosis-related fractures are common in older individuals and post-menopausal women, contributing to morbidity. Post-menopausal women are more likely to suffer through common bone fractures including as vertebral and hip fractures. Osteoporosis is classified into two types: primary and secondary osteoporosis, with up to 30% of post-menopausal women and 50 to 80% of men having factors contributing to osteoporosis. When osteoporosis is age-related and arises in women post-menopause and men in the absence of an underlying disease, it is called primary osteoporosis. When it occurs in the presence of an underlying disease or medication, it is called secondary osteoporosis [1]. Bone density, or bone mineral density (BMD), refers to the concentration of minerals such as calcium and phosphorus in bone tissue, serving as a key measure of bone strength and overall skeletal health. It plays a crucial role in maintaining structural integrity, supporting mobility, and protecting vital organs. High bone density helps prevent fractures and conditions like osteoporosis, while low bone density increases the risk of bone fragility and breaks. Several factors influence bone density, including age, gender, genetics, diet, physical activity, and lifestyle choices. Bone mass typically peaks in early adulthood and gradually declines with age, making proper nutrition and regular weight-bearing exercise essential for maintaining bone strength. Calcium and vitamin D intake, along with healthy lifestyle habits such as avoiding smoking and excessive alcohol consumption, can help preserve bone density. Regular bone health monitoring, including bone density scans, is vital for early detection of potential issues. Prioritizing bone health ensures long-term mobility, reduces the risk of fractures, and enhances overall well-being[2]. Plant-derived phytoestrogens and animal-derived estrogens possess similar chemical structures and may attach to the same estrogen receptors. Phytoestrogenic substances include flavonoids, isoflavonoids, lignans, coumestans, prenylflavonoids, and stilbenoids, primarily from the Leguminosae family, but also from other groups. Certain flavonoids, including flavones, flavonols, and flavanones, exhibit estrogenic characteristics. Phytoestrogens, or xenoestrogens, are plant-derived secondary metabolites that mimic human estrogens in structure and function. These chemicals can operate as agonists or antagonists based on the quantity of endogenous estrogen in the body, without directly affecting biological activities. Phytoestrogens may work through epigenetic mechanisms that are not dependent on estrogen receptors. More than 300 plant species produce these chemicals via the phenylpropanoid pathway, which involves specialized enzymes and results in distinct chemical structures. Phytoestrogens, mostly phenolic substances, consist of isoflavonoids, flavonoids, stilbenes, and lignans. Research shows that phytoestrogens can protect against estrogen-dependent illnesses in both animals and humans. Clinical investigations have demonstrated potential advantages for illnesses including osteoporosis[3]. This review aims to provide a comprehensive understanding of osteoporosis, its classification, causes, and impact, particularly in elderly individuals and post-menopausal women. It will explore the role of bone mineral density (BMD) in maintaining skeletal health and preventing fractures, along with the influence of factors such as age, genetics, diet, and lifestyle. Additionally, the review will examine the potential of phytoestrogens in osteoporosis prevention and management, focusing on their chemical structures, mechanisms of action, and sources. By analyzing clinical and experimental evidence, the review will assess the effectiveness of phytoestrogens in improving bone health and reducing osteoporosis risk, while also considering their broader implications for disease management and overall well-being[4].
Phytoestrogens: Classification And Dietary Sources
Phytoestrogens, also called dietary estrogens and xenoestrogens, are secondary metabolites obtained from plants. Phytoestrogens consist of several chemical scaffolds, including isoflavonoids, flavonoids, chalcones, flavanones, stilbenes, and lignans, with isoflavonoids being the most prominent[5]. Figure 1 illustrates the many classifications of phytoestrogens.
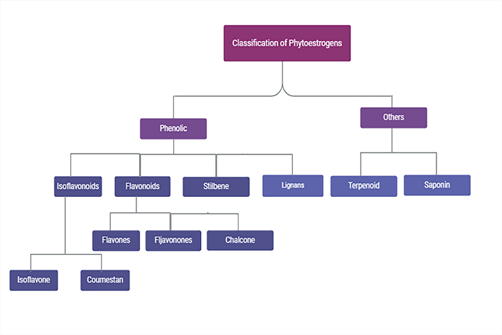
Fig. 1: Classification of Phytoestrogens. Phytoestrogens are composed of various chemical scaffolds, including isoflavonoids, flavonoids, chalcones, flavanones, stilbenes, and lignans, with isoflavonoids being the most significant.
Types of phytoestrogens:
Isoflavones
Estrogens, including 17β-estradiol, estrone, and estriol, are sterane compounds with a hydroxyl group on the third carbon atom of the aromatic ring A and a hydroxyl or oxo group on the carbon atom of the ring D. These sterane chemicals are present in animals. Estrogens and isoflavonoids are structurally related. Phytoestrogenic substances include flavonoids, isoflavonoids, lignans, coumestans, prenylflavonoids, and stilbenoids, primarily from the Leguminosae family but also from other families. Certain flavonoids, including flavones, flavonols, and flavanones, exhibit estrogenic characteristics. The phenylpropanoid metabolic route produces flavonoids by converting phenylalanine to p-coumaroyl-CoA (4-coumarate-CoA) through enzymes such as phenylalanine ammonia-lyase, cinnamate 4-hydroxylase, and 4-coumarate CoA ligase. Chalcone synthase activity causes it to attach to malonyl-CoA, forming the backbone of the flavonoid molecules. These compounds are referred to as chalcones and have two phenyl rings. The enzyme chalcone isomerase closes the ring, forming a three-ring structure. Enzyme activity regulates the metabolic pathway. Isoflavone synthase converts naringenin into flavonols, flavan-3-ols, and other polyphenols (e.g., isoflavonoids), while liquiritigenin converts into genistein, daidzein, and glycitein. Malonyl CoA (3x) and 4-coumarate CoA are converted by stilbene synthase to the stilbene derivative resveratrol. Metabolites of enterodiol, secoisolariciresinol, enterolactone, and matairesinol-type lignans exert estrogenic actions. They are abundant in flaxseed, sesame seeds, and wheat, but also in beans and pumpkins. Green coffee contains daidzin, genistin, daidzein, genistein, formononetin, and biochanin A, as well as three lignans (secoisolariciresinol, matairesinol, and lariciresinol). The milk thistle fruit contains flavonolignan (silychristine, silybinine, and isosilybinin). Plant-derived polyphenols' bioavailability depends on their chemical structure. Lignans with low intestinal absorption are quickly metabolized. Lignans are demethylated and deoxygenated in the gastrointestinal system by the gut flora[3].
Lignans
Plant lignans are polyphenols composed of coniferyl alcohol precursors. Plants contain bi-phenolic chemicals with an oestrogen-like structure, known as phytooestrogens, which have antioxidant qualities. Dietary lignans are found in fiber-rich foods such as whole grains like rye (Secale cereale), oats (Avena sativa), barley (Hordeum vulgare), oilseeds like flax (Linum usitatissimum), sunflower (Helianthus annuus), sesame (Sesamum indicum), cereal, fruits (especially berries), and processed foods[6]. Phytoestrogens include isoflavonoids, flavonoids, stilbenoids, and lignans. Lignans are classified into seven main types: secoisolariciresionol (Seco), pinoresinol (Pino), matairesinol (Mat), medioresinol (Med), sesamin (Ses), syringaresinol (Syr), and lariciresinol. Many lignans exhibit therapeutic effects, including antioxidant, anti-cancer, anti-inflammation, anti-bacterial, and anti-fungal properties . Most bioactivities of lignans arise following their chemical transformation by gut microbiota[7].
Coumestans
Coumestans are a type of plant phenol that have estrogenic action. In 1957, Bickoff and colleagues discovered coumestrol, a phytoestrogen found in ladino clover (Trifolium repens L., Fabaceae), strawberry clover (Trifolium fragiferum L., Fabaceae), and alfalfa/lucerne (Medicago sativa L., Fabaceae). Coumestans having phytoestrogenic activities include coumestrol and 4′-methoxycoumestrol. Coumestrol and genistein have stronger affinity for ERβ than other phytoestrogen substances. Coumestrol has been shown in vitro to prevent bone resorption and promote bone mineralization[8].
Mechanistic Relationship Between Phytoestrogens and Bone Health
Estrogen Receptor Modulation
Estrogens indirectly impact bone metabolism by affecting the synthesis of interleukin 1 and 6 (Il1, Il6), TNFα, and GM-CSFs. They also increase the secretion of IGF-1, TGFβ, and procollagen-1 from osteoblasts. Estrogen insufficiency causes the postmenopausal loss in bone mass and is related with an increased incidence of bone tissue remodelling. Subsequent research have validated the concept that oestrogens play a role in the development of osteoporosis, showing that oestrogen administration reverses bone mass loss caused by ovary removal in premenopausal women. In addition to oestrogen deficiency, other factors that contribute to age-related bone mass loss in women involve secondary hyperparathyroidism, impaired osteoblast function, probably triggered by changes in local cytokine concentrations or systemic growth factors, and deficiencies in certain dietary nutrients, such as vitamin D and calcium[9]. An in situ hybridization assay revealed that ESRRA was regularly expressed in transformation zones during the mouse embryonic development stages and that it could bind directly to the promoter and activate the OPN transcription. Epidemiological research revealed that a single nucleotide polymorphism (SNP) within ESRRA was closely linked to increased bone mineral density (BMD) of premenopausal women. These findings suggested that ERRα might play a role in bone metabolism and osteoblast progenitor development[10]. Plant-based compounds that resemble estrogen are called phytoestrogens. Due to their tissue-specific agonist and antagonist characteristics, their mode of action is similar to that of SERMs. The amount of E2 present may affect their impact. With premenopausal E2 concentrations, isoflavonoids may function as estrogen antagonists; but, with lower E2 concentrations, they exhibit estrogen agonistic activity, which is comparable to postmenopausal women's serum levels. The two main substances that were separated from soybeans are genistein and daidzein, which are members of the isoflavones group. Phytoestrogens have a beneficial effect on osteoporosis because they are agonists in bone tissue. Daidzein has bone-sparing properties against osteoporosis via stimulating the MAPK/PI3K pathways in bones. With its ERβ binding selectivity, a combination of genistein, daidzein, and equol demonstrated neuroprotective effects and enhanced estrogenic processes in the brain while avoiding feminizing effects in the reproductive system. Osteoporosis is defined by weakening bone microstructures with total bone loss and an increased risk of fractures. It can be linked to a number of events, including age and menopause. Osteoporosis-related fractures have occurred or will occur in every other postmenopausal woman. In actuality, the first stage of postmenopausal osteoporosis (PMOP), which lasts for three to five years and is marked by a rapid loss of trabecular bone structure, may be associated with E2 deficiency. In contrast, the second stage, which lasts for ten to twenty years and affects both men and women equally, involves an age-dependent loss of both cortical and trabecular bone structure[11]. E2 controls the osteoprotegerin (OPG), RANK, and receptor activator NF-κB (RANK) ligand (RANKL) pathways. In fact, the biology of osteoclasts depends on these three chemicals, which are not redundant. While OPG-deficient mice show excessive bone resorption and severe osteoporosis, RANK- or RANKL-deficient mice have no osteoclasts at all, resulting in osteopetrosis. The primary impact of E2 is the inhibition of osteoblasts' RANKL expression, which lowers osteoclast activation. OPG is a natural antagonist of RANKL, and its increased expression aids in this process[12].
Regulation of Osteoblast and Osteoclast Activity
The intricate process of bone metabolism is influenced by a number of cytokines, the balance of the RANKL/RANK/OPG pathway, and osteoblastic and osteoclastic activity (Figure 2). Like other TNF family receptors, the receptor activator of nuclear factor κB is incapable of activating innate protein kinase. As a result, adaptor molecules are required to connect to the RANK intracytoplasmic domain in order to activate signaling. TNF receptor-associated factors (TRAFs) bind to specific locations in the RANK cytoplasmic domain and regulate downstream molecules. Trafs binding to RANK motifs (Motif-1, -2, or -3) on the RANK cytoplasmic domain is a key step in the mediation of downstream signaling after ligation of RANKL to RANK. After NF-κB is activated by motif-1, the NF-κB intracellular signaling cascade is subsequently promoted. The last stage of RANK activation is the translocation of NF-κB to the nucleus, which controls the production of several genes, including c-Fos, the nuclear factor of activated T cells 1, and several bone morphogenetic proteins (BMPs)[13]. RANKL, a crucial regulator of osteoclastogenesis, promotes and increases bone loss and osteoclastogenesis; hence, animals lacking Due to a lack of osteoclasts, RANKL had osteoporosis. When RANK is activated in osteoclasts by RANKL, which was generated by osteoblasts, osteoclast maturation occurs. By secreting acid and lytic enzymes (such as tartrate-resistant acid phosphatase and cathepsin K), mature osteoclasts stick to the surface of the bone and encourage bone resorption. A decoy receptor belonging to the TNF receptor family, osteoprotegerin reduces osteoclastogenesis by blocking RANK activation by RANKL. OPG expression and synthesis are regulated by a number of cytokines, including transforming growth factor β (TGF-β), BMPs, steroid hormones (17β estradiol), and interleukins 1 α, 18 α, and TNF-α. BMD rapidly declines after menopause, which causes osteoporosis. This happens as a result of aging-induced decreases in estrogen release and decreased intestinal mineral absorption. The effects of estrogen may be mimicked by soy isoflavones. The estrogenic actions of daidzein can cause an improvement in bone resorption. Supplementing with soy isoflavones can reduce the risk of fractures in postmenopausal women. Isoflavones prevent bone loss in ovariectomized (OVX) rats. ER α and ER β are among the estrogen receptors that soy isoflavones bind to. They preferentially attach to ER β. It proves that soy isoflavones function as specific estrogen receptor modulators[14].
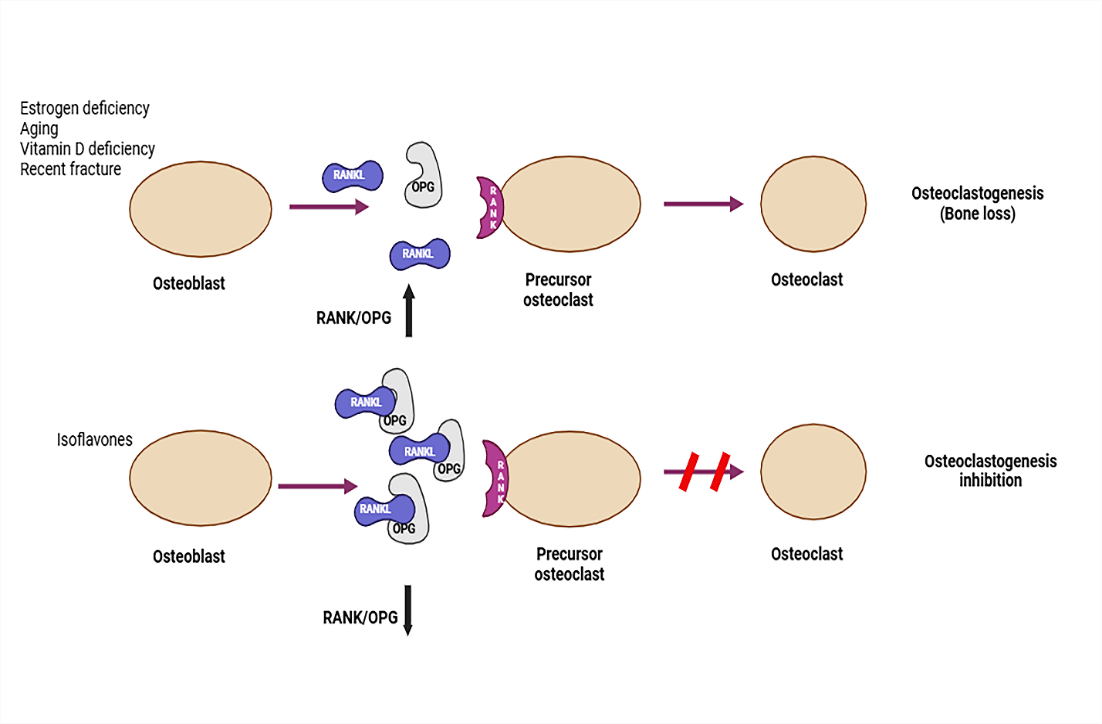
Fig. 2: Regulation of Osteoblast and Osteoclast Activity. Osteoporotic fractures are linked to aging, estrogen deficit, vitamin D deficiency, and prior fractures. Bone homeostasis relies on the OPG/RANKL balance. RANKL activates osteoclasts, promoting bone resorption. Soy isoflavones lower the RANKL/OPG ratio, inhibiting osteoclastogenesis and fostering bone formation.
Antioxidant and Anti-inflammatory Effects
Reactive oxygen species (ROS) are molecules and free radicals that mostly arise as byproducts of electrons that have escaped from the mitochondrial electron transport chain throughout aerobic respiration. Examples of these include superoxide anion, hydrogen peroxide, and hydroxyl radical. ROS are essential for controlling cellular activities such as proliferation, survival, metabolism, apoptosis, and differentiation. ROS are interesting molecules in bone regeneration because they can serve several functions. Their usefulness stems from their ability to operate as an intracellular marking agent, which is necessary for cell signal transmission. However, when ROS levels raise owing to aging or inflammation, they can cause cellular imbalance in reduction-oxidation pathways, resulting in oxidative stress. Oxidative stress alters essential cellular physiological responses through signal transduction, transcription factors, and ncRNAs, causing nuclear and mitochondrial DNA damage and beginning DNA repair pathways. Chronic oxidative stress has been shown to cause disease by destroying bone and killing cells[15].
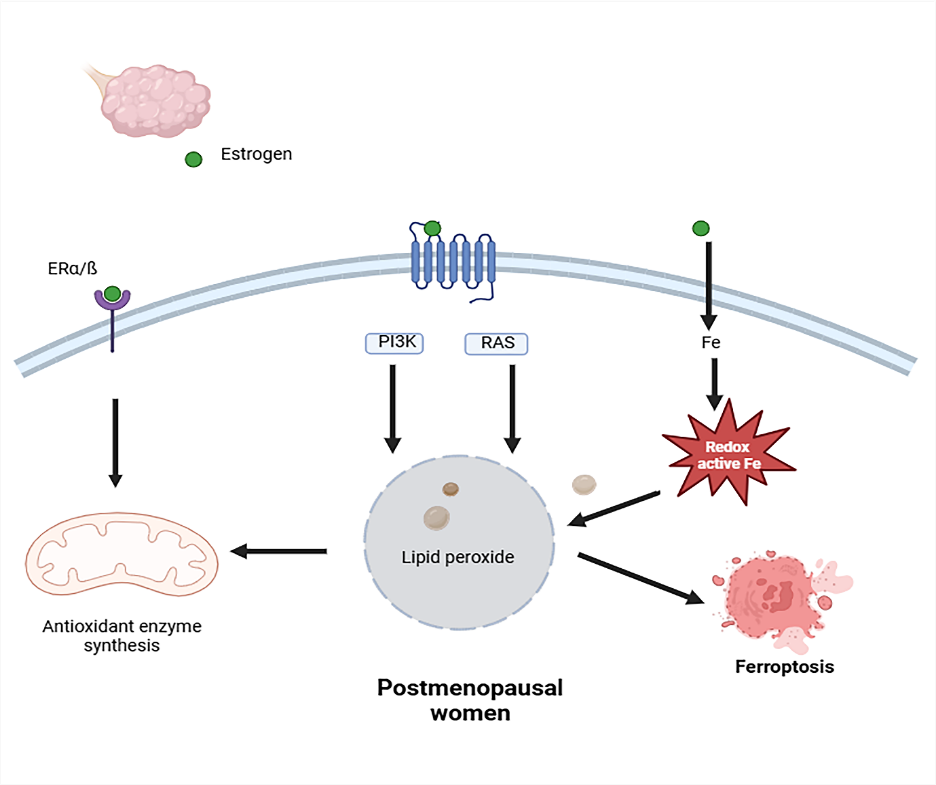
Fig. 3: Oxidative stress in a high-risk osteoporotic population. postmenopausal women. Estrogen receptors and G-protein-coupled estrogen receptor-1 trigger an intracellular antioxidant response. Estrogen insufficiency caused lipid peroxide buildup and promoted ferroptosis.
ROS induces enhanced expression of osteoclastogenic markers, including c-Fos, NFATc1, and TRAP. GSH metabolism and ferroptosis play a role in ROS-mediated control of bone metabolism. The sulfhydryl group (-SH) in GSH reduces hydrogen, gives the free radical a paired electron, and reduces its oxidative and aggressive nature. After connecting with reducing hydrogen, oxidized GSH with a disulfide link transforms into reduced glutathione, inhibiting further oxidation. GSH enhances osteogenic differentiation by boosting the expression of Collagen I, OCN, and ALP. Elevating GSH suppresses RANKL-mediated osteoclastogenesis via inhibiting the ROS/NF-κB signaling pathway (Figure 3) [16].
Calcium Homeostasis and Bone Mineralization
The body produces parathyroid hormone (PTH), a hypercalcemic hormone that increases plasma calcium via acting on the kidney, bone, and/or small intestine when plasma calcium levels fall. The parathyroid glands release PTH largely through their primary cells. The polypeptide has 84 amino acids. Calcitonin, another hormone, opposes its activity.
The parathyroid hormone-related protein (PTHrP) is a protein from the parathyroid hormone family. PTHrP functions as an endocrine, autocrine, paracrine, and intracrine hormone. It governs endochondral bone development by keeping the growth plate at a consistent width. The protein is secreted by mesenchymal stem cells. Cancer cells, especially breast and squamous-cell lung carcinoma, may leak it on occasion[17].
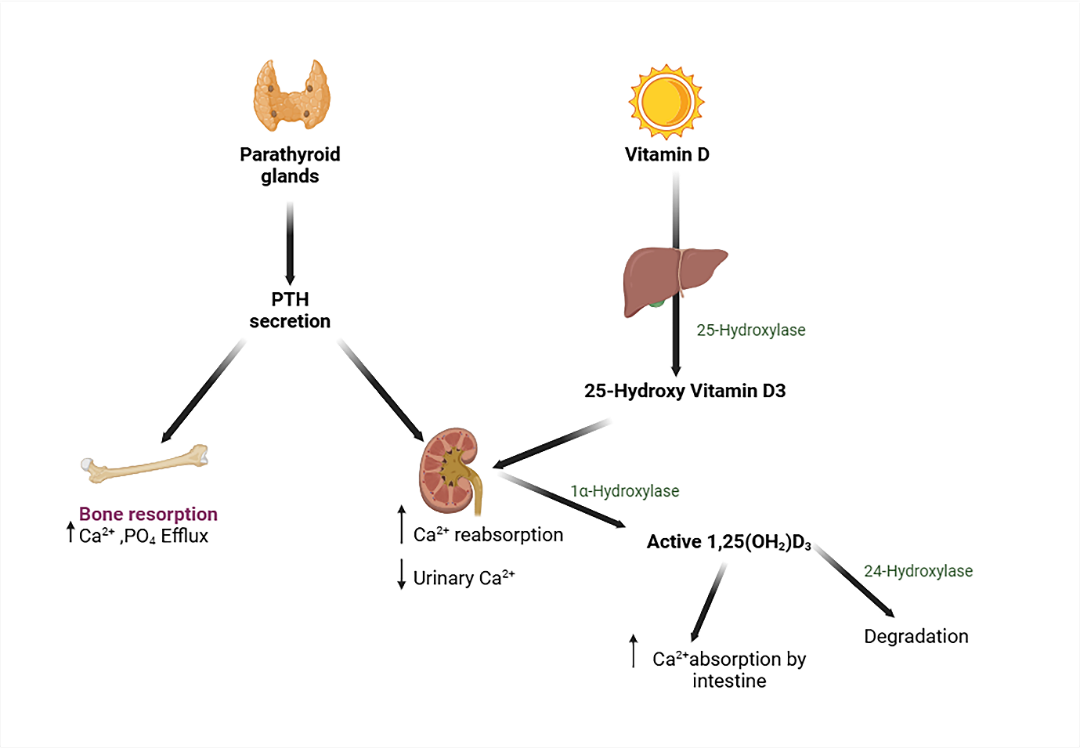
Fig. 4: Calcium regulation at the axis of the parathyroid, bone, kidney, and gut
Serum calcium levels are recognized by the calcium-sensing receptor (CaSR) on parathyroid cell surfaces. CaSR signaling suppresses PTH secretion. This inhibition is alleviated when serum calcium or magnesium levels are low, allowing for PTH secretion. PTH stimulates bone resorption and calcium release, whereas kidneys reduce calcium excretion. PTH1R stimulates NaPi-IIa internalization in kidney proximal tubules, reducing phosphate absorption and altering CYP27B1 and CYP24A1 expression, leading to 1,25(OH)2D3 production. Vitamin D is activated through hydroxylation by 25-hydroxylase (CYP2R1) in the liver and 1α-hydroxylase (CYP27B1) in the kidney, resulting in active 1,25(OH)2D3 that binds to the vitamin D receptor. Inactivation requires 24-hydroxylase (CYP24A1). FGF23 is released by osteocytes and binds to the FGFR-klotho complex in kidney proximal tubules. This lowers PTH transcription and plasma membrane expression of NaPi-IIa and NaPi-IIc, leading to increased phosphate excretion. FGF23 inhibits CYP27B1 and stimulates CYP24A1, resulting in decreased 1,25(OH)2D production (Figure 4) [18].
Evidence From Preclinical and Clinical Studies
Preclinical Studies: Effects on bone mineral density (BMD) in osteoporosis models
The GCs utilized in GIOP mouse models include methylprednisolone (MP) (14%) in rats, prednisolone (47%), MP (24%), and DEX (12%) in mice, and dexamethasone (DEX) (49%), and prednisone (22%). In rats, the predominant routes of administration were subcutaneous (SC) (45%), intramuscular (IM) (22%), and via oral (22%). The intraperitoneal (IP) and intravenous (IV) methods were less popular. In mice, the most common method for SC was by surgically placed pellets (pellet in subcutaneous: PSC) (61%). Other methods of administration were less commonly employed (16% for P.O. and 16% for SC). In rabbits and lambs, the methods of administration were intramuscular and subcutaneous. DEX was supplied to rats at several doses, from 0.1 mg/kg daily for 60 days to 25 mg/kg twice each week for six weeks. Prednisone was given in doses of 1.5 mg/kg per day for 90 days and 6 mg/kg per day for 90 weeks. Prednisolone was delivered to mice at varying doses of 0.8 mg/kg per day for three weeks and 4 mg/kg per day for three weeks. In rabbits and lambs, DEX was given at 3 mg/kg twice weekly for 12 weeks and prednisolone at 0.6 mg/kg five times weekly for seven months, respectively[19]. (Table 1)
Table 1: Preclinical studies to evaluate the effect of Phytoestrogen
|
Phytoestrogen |
Model (method) |
Species |
Age / Weight (g) |
Duration |
Outcome |
References |
|
20 mg/kg psoralen |
OVX |
Female C57BL/6 mice |
7? 10 week / 20 g |
21 days |
Promote callus formation and inhibit osteoclast genesis |
Kui Huang et al.[20] |
|
Fisetin 2.5 mg/kg |
OVX |
Sprague-dawley rat |
3 months / 200-230 g |
8 weeks |
Reduces osteoclast activity |
Peng Feng et al.[21] |
|
100 mg/kg kaempferol |
OVX |
Female SD rats |
three months / 220 g |
16 weeks |
Increased osteogenic differentiation of cultured osteoblasts |
Chirag Kulkarni et al.[22] |
|
Pueraria tuberosa (200 mg/kg) |
OVX |
Wistar rats |
NR / 220–250 g |
90 days |
Exhibited antiosteoporotic activity |
Swaha Satpathy et al.[23] |
|
Trifolium pratense (500 mg/kg/day) |
OVX |
Female Sprague–Dawley rats |
8-weeks / 190–230 g |
NR |
Inhibited bone resorption |
Yixian Quah et al.[24] |
|
Panax ginseng powder (6.2g), Wolfiporia extensa (12.4g), and honey (41.9g),Rehmannia glutinosa (39.9g), P. lobata |
OVX |
Sprague (SD) rats |
9 weeks / 170–175g |
12 weeks |
Synergistic effects |
Kim, M., et al.[25] |
|
Matricaria chamomilla Linn (400mg/kg) |
methyl prednisolone (28 mg/kg) induced osteoporosis |
Wistar rats |
5–6 months / 200–220 g |
8 weeks |
Potential therapeutic benefits |
Raja, A., et al.[26] |
|
chrysin (100 mg/kg) , naringenin (100 mg/kg) |
Isotretinoin induced osteoporosis |
Y59 female rats |
3 months / NR |
14 days |
Lower efficacy compared with alendronate |
Oršoli?, N., et al.[27] |
|
Cissus quadrangularis (500 mg/kg/day) |
OVX |
Female Balb/c mice |
8–10 weeks / 23g |
45 days |
Suppresses the RANKL-induced osteoclastogenesis |
Azam, Z., et al.[28] |
|
Epimedium brevicornum, Cortex eucommiae, Malaytea scurfpea, Semen cuseutae ( 3.0 g/kg/d) |
OVX |
Sprawl-Dawley rat |
NR / 200 ± 20 g |
|
|
|
OVX – ovariectomy, NR – not reported
Human Studies:
Clinical studies on the efficacy of estrogen in osteoporosis treatment have produced promising results. The Women's Health Initiative (WHI) has found that estrogen therapy may help postmenopausal women maintain bone density and minimize their risk of fractures. The beginning of estrogen therapy near menopause appears to provide the most effective protection against osteoporosis-related fractures. Meta-analyses and observational studies that show estrogen therapy improves bone mineral density and lowers the incidence of fractures offer more evidence in favor of these conclusions. In summary, these results imply that estrogen therapy might be a useful strategy for preserving bone health and averting difficulties from osteoporosis in women going through menopause[30].( Table 2)
Table 2: Clinical studies to evaluate the effect of Phytoestrogen
|
Phytoestrogen |
Study design |
Participants, age, sex |
Dose / Duration |
Outcome |
Reference |
|
Equol and resveratrol |
Randomized control trial |
50 - 55 years |
75mg twice daily |
Improving bone density formation |
Corbi, G., et al. (2023)[31] |
|
100 mg isoflavones, 520 mg black cohosh, 400 mg chasteberry, and 500 mg evening primrose oil |
Randomized |
45–60 years |
12 weeks |
Improve menopausal symptoms and decrease inflammation markers |
Rattanatantikul, T., et al. (2022)[32] |
|
Plant-based diets |
Case-control study |
45–65 years |
NR |
Protective effect in preventing bone loss |
Ghadiri, M., et al. (2023)[33] |
|
Cornus mas extract |
Interventional double-blind placebo-controlled randomized study |
45–60 years |
8 weeks |
No beneficial effect on OC and TC |
Aryaeian, N., et al. (2021)[34] |
|
Ginger (Z. Officinale), curcumin (C. Longa L.), |
Prospective, factorial, randomised, triple-blinded, placebo-controlled study |
Postmenopausal women |
4-month |
Improved serum osteocalcin, ALP, hs-CRP, and SOD levels |
Salekzamani, Y., et al. (2023)[35] |
|
PMA-zeolite-clinoptilolite |
Randomized |
56 to 74 years |
12 months |
Potential |
Kraljevi? Paveli?, S., et al. (2021)[36] |
|
Osteo-f |
Randomized, double-blind, placebo-controlled clinical trial |
45 years |
24 weeks |
Anabolic effects |
Kim, M.H., et al. (2023)[37] |
|
R. Coreanus miq. And a.Membranaceus bunge extract |
Randomized |
females with postmenopausal syndrome |
12 weeks |
Improved the menopausal symptoms |
Park, J.-S., et al. (2022)[38] |
OC – osteocalcin, TC - C-terminal telopeptide, ALP - Alkaline phosphatase , hs-CRP - high-sensitivity C-reactive protein , SOD - Superoxide dismutases
Potential Clinical Applications
Phytoestrogens as an alternative to hormone replacement therapy (HRT)
HRT is typically composed of two hormones: oestrogen and progesterone.
Oestrogen is taken to treat short-term, intermediate-term, or long-term problems caused by low hormone levels. Giving progesterone for 10-12 days every month can avoid endometrial hyperplasia and carcinoma. Oestrogen causes the uterine lining to thicken. After an abortion, a woman only has to undergo estrogen replacement medication. Hormone replacement treatment (HRT) is recommended for oestrogen deficiency and can be administered orally, transdermally, implanted, or locally. HRT can be administered using one of three different regimens. Oestrogen is administered daily for women under 54 who still have periods, and progestogen for the last stages of the menstrual cycle. This allows the endometrium to fall, which usually causes a withdrawal bleed. Although breakthrough bleeding is possible, several treatments can reduce bleeding to every three months. Traditional hormone replacement therapy (HRT) includes estrogens and synthetic progesterone to address menopausal symptoms. Menopause in women is connected with several difficulties, including hot flushes, memory, vaginal dryness, and osteoporosis. Studies show that phytoestrogen ingestion helps lower menopausal symptoms in women. The addition of soy isoflavones to the diet reduced menopausal flushes in women. Lower doses of genistein increased bone mineral density, whereas higher doses produced no improvement in ovariectomized rats. In humans, consuming phytoestrogens improves memory. In general, menopausal difficulties have not been noticed in women who eat a balanced diet that includes plant-based foods[39].
Function of phytoestrogens in reproduction
Phytoestrogen increases animal ovarian progesterone and cyclic adenosine monophosphate synthesis. Exogenous estrogen-like chemicals also have an effect on oocyte maturation throughout the zygote's development in the preimplantation stage. Green tea and Indian turmeric contain phytoestrogens, which limit cell proliferation and promote apoptosis. Consuming isoflavones has been linked to changes in pituitary function and hypothalamus activity. Combining isoflavones and lignans can suppress hot flashes, which are common menopausal symptoms for women[40].
Phytoestrogen impacts on the immune system
Genistein has been shown to have a considerable influence on the suppression of antigen-specific immunological responses in vivo, as well as lymphocyte proliferation in an in vitro research model. In addition to cytokine production, genistein stimulates T cell cytotoxicity[41].
Impact of phytoestrogens on osteoporosis
A reduced estrogen levels in the body causes bone turnover, which can be enhanced by bone reabsorption. Osteoclast activity confirms the increased bone resorption. Gonadal dysfunction causes epiphyseal growth plates to close slowly. Several phytoestrogens, including genistein and daidzein, have been proven in rats to reduce bone loss when combined with coumetarol. The success of these treatments is determined by a variety of parameters, including the phytoestrogen dose, duration of action, and animal model used. The chemical equol, derived from daidzein, is essential for the bone-sparing effects. Some folks create equol from gut flora and gain more from soy isoflavone compared to others[42].
Natural phytoestrogens as neuroprotective agents for Parkinson's illness
Phytoestrogens have neuroprotective benefits on Parkinson's disease (PD) via activating three signaling pathways: ESα, ESβ, and GPER. Adding phytoestrogens to PD improves neural development and neuronal survival[43].
Significance of phytoestrogens in cancer
Genistein, a phytoestrogen, has been studied for its potential anticancer effects by modulating cell cycle and apoptosis. It may inhibit NF-kB and Akt signaling pathways. These signaling pathways are crucial for the regulation of homeostasis and cell survival, as is apoptosis. Phytoestrogen was applied to block angiogenesis and metastasis processes by using its antioxidant potential, thus preventing the basic damage that occurs during cancer[44].
Future Directions and Research GAPS
The human body receives biologically active substances from a diet high in raw fruits and vegetables, such as phytoestrogens, which can function as agonist or antagonist estrogenogenic molecules, antioxidants, and minerals that reduce inflammation. Thus, they can trigger protective mechanisms. Consuming phytoestrogens at multiple meals throughout the day might lead to long-term accumulation and either positive or negative consequences on health. The ideal dose, manner of administration, and frequency of usage of dietary phytoestrogens to achieve therapeutic benefits remain unclear. The number of osteoporosis patients is expected to rise in step with the senior population, as is the number of people suffering from fractures of fragility such as hip fractures. More extensive bone evaluations may be possible now or in the near future. HR-pQCT is helpful in assessing bone microstructure and can clearly illustrate how postmenopausal osteoporosis patients' bones react differently to various osteoporosis medications[45]. Vitamin D supplementation was linked to decreased rates of gastrointestinal adverse reactions and mortality in postmenopausal women undergoing antiresorptive osteoporosis medication. Calcium supplementation had no effect on any of the endpoints of interest. Vitamin D supplementation appears to be more beneficial than calcium supplements alone due to the relationship between calcium absorption and vitamin D levels. Romosozumab efficiently raises BMD and lowers fracture risk, especially when given as an initial therapy in high-fracture-risk individuals. Sequential therapy combined with antiresorptive treatment maximizes long-term benefits. While typically well accepted, its cardiovascular safety profile necessitates additional long-term research to ensure its safety in clinical practice. Additional research is needed to prove efficacy and safety in secondary osteoporosis patients. Common anti-osteoporosis treatments include Teriparatide, Abaloparatide, Alendronate, Raloxifene, Denosumab, and Romosozumab. The above monotherapies may improve bone mineral density and reduce fracture risk in osteoporosis patients to varied degrees. While anti-osteoporosis medications have made great progress in prevention and treatment, there is currently an obstacle in the discovery of new therapeutic treatments[46]. Phytoestrogens, which include isoflavones, lignans, and coumestans, exhibit both estrogenic and antiestrogenic properties. Plant foods containing these chemicals include soybeans, flaxseeds, chickpeas, and lentils. Phytoestrogens are increasingly being studied for their possible health advantages, notably in kidney protection. Phytoestrogens have been proven to lower oxidative stress and inflammation in the kidneys, both of which contribute to the development of renal disease[47].
CONCLUSION:
Phytoestrogens may protect bones by reducing bone resorption while boosting bone growth. Phytoestrogens, particularly isoflavones, may protect against alveolar bone loss in postmenopausal osteoporosis patients. Well-designed clinical trials are required to determine the therapeutic potential of phytoestrogens on skeletal health, particularly in postmenopausal women. Phytoestrogens have the potential to be developed as cost-effective preventive and therapeutic techniques in the treatment and prevention of bone loss in postmenopausal osteoporosis. Elaborative studies are needed to confirm the efficacy of isoflavones on the RANKL/RANK/OPG pathway. Identifying the effects of soy isoflavones on the RANKL/RANK/OPG pathway can guide future research and lead to effective treatment strategies for bone disorders. A plant-based diet rich in certain antioxidants may enhance bone mineral density, lower the incidence of osteoporosis and age-related bone loss, and help young people reach their ideal bone mass, according to the evaluated studies. As a result, natural antioxidants have been advocated as a means of preventing OS bone deterioration, with a special focus on antifracture therapy. To achieve this, more research should be done to elucidate the molecular processes that underlie osteocyte activity and how they relate to the intracellular oxidative state.
REFERENCES


Sudhakar Pachiappan, Gayathiri Muthusamy, Akshaya Sivakumar*, Exploring the Effect of Phytoestrogens on Bone Density and Osteoporosis Prevention, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 7, 1719-1735. https://doi.org/10.5281/zenodo.15876389
 10.5281/zenodo.15876389
10.5281/zenodo.15876389
