Dr. C. S. N Institute of Pharmacy.
The successful lab-grown development of a human spine would represent a groundbreaking achievement in regenerative medicine and biological sciences. Such a breakthrough could revolutionize treatments for spinal injuries, degenerative diseases, and congenital spine disorders.
Scientists have officially grown a notochord—the tissues that act as the “GPS for the developing embryo” by guiding the formation of the spine and nervous system “What’s particularly exciting is that the notochord in our lab-grown structures appears to function similarly to how it would in a developing embryo. It sends out chemical signals that help organize surrounding tissue, just as it would during typical development.”
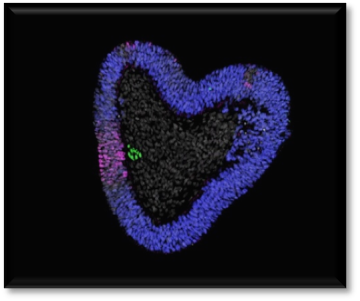
Section of a human trunk-organoid. The notochord (green) causes neural tissue (in blue) to be patterned (in magenta). Cell nuclei stained in white. Credit: Tiago Rito Previous attempts to create a notochord from human stem cells have failed because scientists didn’t have a firm grasp on the ideal timing of the introduction of certain biological “ingredients” to spur growth.
Now that they have grown a notochord—albeit a simple one—the researchers hope this lab-grown spinal tissue will give us insights into spine-related birth defects and other intervertebral discs (the cushy tissue that lies between our vertebrae).
The formation of the vertebrate body involves the coordinated production of trunk tissues from progenitors located in the posterior of the embryo. Although in vitro models using pluripotent stem cells replicate aspects of this process1,2,3,4,5,6,7,8,9,10, they lack crucial components, most notably the notochord—a defining feature of chordates that patterns surrounding tissues11. Consequently, cell types dependent on notochord signals are absent from current models of human trunk formation. Here we performed single-cell transcriptomic analysis of chick embryos to map molecularly distinct progenitor populations and their spatial organization. Guided by this map, they was investigated how differentiating human pluripotent stem cells develop a stereotypical spatial organization of trunk cell types. They found that YAP inactivation in conjunction with FGF-mediated MAPK signalling facilitated WNT pathway activation and induced expression of TBXT (also known as BRA). In addition, timely inhibition of WNT-induced NODAL and BMP signalling regulated the proportions of different tissue types, including notochordal cells. This enabled us to create a three-dimensional model of human trunk development that undergoes morphogenetic movements, producing elongated structures with a notochord and ventral neural and mesodermal tissues. Our findings provide insights into the mechanisms underlying vertebrate notochord formation and establish a more comprehensive in vitro model of human trunk development. This paves the way for future studies of tissue patterning in a physiologically relevant environment.
Main
The formation of the vertebrate body axis is an evolutionarily conserved process that requires coordinated generation of multiple cell types from a population of progenitors in the caudal embryo12. The node, a midline structure, has a central role in this process13. It secretes factors to organize forming trunk tissues14 and later gives rise to the notochord15,16, the mesodermal rod that provides mechanical and signalling cues to the embryo, and the floor plate, the ventral midline domain of the neural tube that patterns neural tissue. Various signalling pathways, including WNT, BMP, NODAL and FGF17, have been implicated in this process, but how these produce and organize the cell types necessary to form the body is not clear.
In vitro models of stem cell differentiation aimed to mimic signalling conditions around the node have been developed including gastruloids1,3,4,5, axially elongating organoids9,10, trunk-like structures2, spinal cord organoids6, somitoids7, and axioloids8. Although these models generate varying amounts of neural, endodermal and mesodermal tissue, they often lack key components, most notably the notochord and its dependent tissues such as the floor plate. This raises the possibility that notochord specification has specific signalling requirements not met in current models.
Anatomy and physiology of the spinal cord
Extending to nearly 45 cm in humans, the spinal cord is part of the central nervous system (CNS), starting at the foramen magnum and reaching the extremities of the first lumbar vertebral. From outside to inside, the spinal cord is concealed under the three meninge membranes (dura mater, arachnoid mater, and pia mater) and is secured by the bony compositions of the spinal column. The vertebral (spinal) column is itself comprised of the cervical, thoracic, lumbar, and sacrococcygeal segments[6],[7], which together form the central axis of the skeleton of vertebrates. The spinal cord is nourished via three main blood arteries oriented vertically. However, the distribution of blood is not quite the same among different segments, wherein the average blood flow in the cervical and lumbar segments is higher than 40% as opposed to the thoracic segment, albeit spinal veins follow along an identical course to that of the arteries. The spinal cord parenchyma is shielded by a blood-spinal cord barrier (BSCB) [7],[8],[9]. The main constituents of the spinal cord are gray and white matter, with the latter surrounding the former. Gray matter, also known as the H-shaped region, contains the cell bodies of motor neurons and interneurons plus glial cells and blood vessels, while white matter contains glial cells and nerve fibers of the ascending and descending tracts. These glial cells are made of oligodendrocytes, astrocytes, and microglia [8], [9],[10], that the first two are originated from the neuroectoderm, and the last derived from blood monocytes [6]. The oligodendrocytes produce myelin within the CNS, while astrocytes act to direct the migration of neurons during embryonic development. However, in a mature CNS, astrocytes constitute the structural scaffolding for other cells. Astrocytic foot processes are a prime contributor to the formation of the blood-brain barrier and shelter the CNS from external impacts at the pial surface [6]. Cells in a normal spinal cord are enclosed by an extracellular matrix (ECM), including laminin, fibronectin, tenascin-C/-R, proteoglycans (such like chondroitin sulfate proteoglycans (CSPGs) and heparan sulfate proteoglycans (HSPGs), thrombospondin, hyaluronan, semaphorins, growth factors, and other proteins[11]. The structural frame of the spinal cord parenchyma is essentially dependent on the interworking of neurons and glial cells with the surrounding ECM, which also contributes to the regulation of both cellular and biochemical processes. As a result, the survival, migration, proliferation, differentiation, neurite extension, and synaptogenesis of cells rely heavily on the interaction between glia and ECM [11].
Complexity of growing human spine
Your spine is a complex structure of small bones, cushioning disks, nerves, joints, ligaments and muscles. This part of your anatomy is at risk of injury, arthritis, herniated disks, pinched nerves and other conditions.
Human trunk development, including spine and spinal cord organogenesis, is a coordinated, orderly, and interdependent process with spatiotemporal tissue patterning. However, the underlying cellular and molecular mechanisms remain largely unclear due to the lack of an effective model that can simulate the early development of human body axis. Here, we reported the long-term patterning and dynamic morphogenesis of human trunk through the formation of spine-spinal cord organoids (SSCOs) self-organized from three-dimensional culture of human PSC-derived neuromesodermal progenitors (NMPs). The SSCOs resembled the morphogenetic features of spine and spinal cord along the anterior–posterior axis, and showed the chondro-osteogenic and neural trajectories consistent with developmental dynamics of spine and spinal cord in gestational embryo through single-cell RNA sequencing (scRNA-seq). In addition, we identified a new HMMR+ bipotent cell population with self-renewal ability and neural/mesodermal competence but distinct from NMPs, which may be involved in trunk development and represent an invaluable tool for disease modeling of spine- and spinal cord-related disorders.
Advancements in Lab-Grown Human Trunk Models
The stem cells formed a miniature ‘trunk-like’ structure, which spontaneously elongated to 1-2 millimeters in length. It contained developing neural tissue and bone stem cells, arranged in a pattern that mirrors development in human embryos. This suggested that the notochord was encouraging cells to become the right type of tissue at the right place at the right time.
The scientists believe this work could help to study birth defects affecting the spine and spinal cord. It could also provide insight into conditions affecting the intervertebral discs – the shock-absorbing cushions between vertebrae that develop from the notochord. These discs can cause back pain when they degenerate with age.
Cell-based treatments for SCI regeneration
Cell therapy, a revolutionary strategy, offers a hopeful promise to treat various types of diseases, even intractable ones[47],[48],[49]. This strategy involves two approaches. The one that first comes to mind is transplanting exogenous cells aiming to regenerate the injured site or to impose supportive effects on survived cells. The next one is directing or enhancing the functions of endogenous progenitor cells[50],[51],[52]. The positive effects of this therapeutic method, depending on the type of cell, the timing, and the route of administration include modulating the immune system, replacing the lost neurons, suppling vital neurotrophic factors and ECM components, and subsequently encouraging regeneration[53],[54],[55].
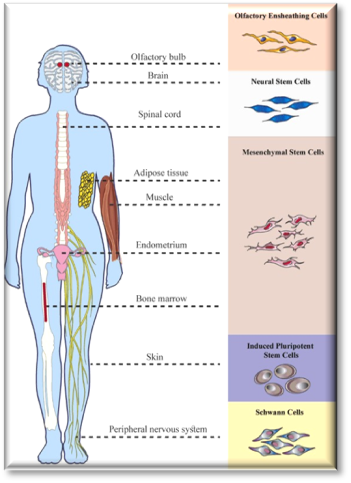
There are many studies on the efficacy of different cell types in pre-clinical and clinical SCI trials[56],[57]. Researchers have studied various adult and stem cell types, which are as follows: neural stem cells (NSCs), embryonic stem cells (ESCs), mesenchymal stem cells (MSCs), induced pluripotent stem cells (iPSC), oligodendrocyte precursor cells, endothelial progenitor cells, olfactory ensheathing cells (OECs), Schwann cells, amniotic epithelial cells, keratinocytes, mast cells, mononuclear cells, T cells, and also genetically modified cells[58],[59],[60]. In this paper, to avoid prolonging the content, and meanwhile, to review thoroughly, we will focus on the cell types that have been studied the most and are of higher promise (Fig. 3)
Paving the way for clinical applications
The researchers’ models open new doors for understanding developmental conditions of the spine, such as congenital scoliosis, as well as diseases involving tissues that arise from the same region of the embryo, known as the paraxial mesoderm. These include skeletal muscle and brown fat in the entire body, as well as bones, skin and lining of blood vessels in the trunk and back. hopes that researchers will be able to use the new stem cell models to generate differentiated tissue for research and clinical applications, such as skeletal muscle cells to study muscular dystrophy and brown fat cells to study type 2 diabetes. Such work would provide a foundation for devising new treatments.
Notes
These structures are simplified models of the body which contain only a small number of cell types. They develop for just a few days and cannot form embryos. Their main purpose is to study aspects of human development that have been difficult or impossible to investigate directly.
REFERENCES


Shaik Parveen*, Dundi Harika, Nandam Deepak Japhanya, Ganta Yamini, Complexity of In - Vitro Cultivation of Human Spine by Researchers, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 3, 2193-2198. https://doi.org/10.5281/zenodo.15078552
 10.5281/zenodo.15078552
10.5281/zenodo.15078552
