S. N. D. College of Pharmacy, Babhulgaon
The human gastrointestinal tract harbors a complex microbial ecosystem that profoundly influences host metabolism, immunity, and overall homeostasis. Conventional probiotics have long been employed to support gut health; however, their limited specificity and passive functionality restrict their therapeutic scope. The advent of CRISPR-Cas genome engineering has enabled the creation of smart probiotics—live, programmable microorganisms capable of sensing pathological cues and autonomously delivering therapeutic molecules at disease sites. This review explores the design principles, molecular mechanisms, and therapeutic potential of CRISPR-engineered smart probiotics. Emphasis is placed on the integration of CRISPR-Cas systems with synthetic gene circuits, biosafety mechanisms such as kill-switches, and regulatory elements that enable precise spatiotemporal control of therapeutic output. Applications span metabolic disorders, inflammatory and infectious diseases, cancer immunotherapy, and precision microbiome modulation. Furthermore, the paper addresses key challenges, ethical concerns, and regulatory barriers associated with the clinical translation of genetically modified live biotherapeutics. Overall, CRISPR-engineered smart probiotics represent a transformative convergence of synthetic biology and microbiome science, offering a path toward personalized, adaptive, and safer therapeutic interventions.
The human gastrointestinal (GI) tract hosts a remarkably dense and diverse microbial ecosystem that is pivotal in regulating host physiology, immune function, and metabolic homeostasis. Within this complex environment, probiotics—defined as live microorganisms conferring a health benefit when administered in adequate amounts—have long been a focus for restoring microbial balance and supporting gut health. However, recent scientific advancements, particularly the integration of synthetic biology and precise gene editing, notably CRISPR-Cas systems, are dramatically reshaping the field, reimagining probiotics as sophisticated, programmable biotherapeutics [1, 2].
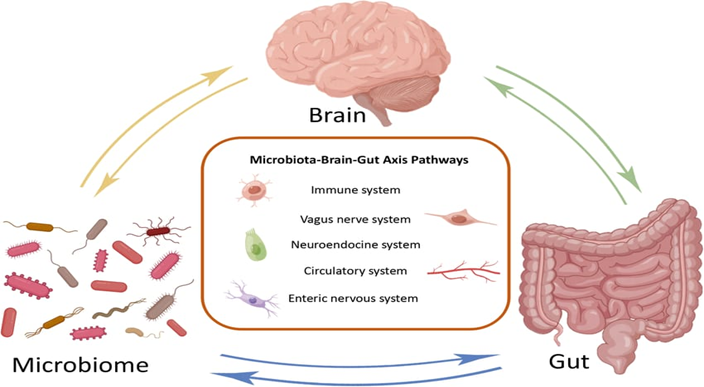
Figure-1-Bidirectional communication pathways in the Microbiota–Gut–Brain axis. Reproduced with permission from Ref [10] Yuan, C., He, Y., Xie, K., Feng, L., Gao, S., & Cai, L. (2023). Review of microbiota gut brain axis and innate immunity in inflammatory and infective diseases. Frontiers in Cellular and Infection Microbiology, 13, 1282431
Recent breakthroughs in microbiome research have illuminated the profound systemic influence of gut microbes. Dysbiosis, or the disruption of this microbial equilibrium, is now implicated in a broad spectrum of disorders, including inflammatory bowel disease (IBD), metabolic syndrome, cancer, and various autoimmune conditions [3, 4]. This fundamental understanding has spurred the development of Next-Generation Probiotics (NGPs) and Live Biotherapeutic Products (LBPs). Moving beyond traditional Lactobacillus and Bifidobacterium strains, these engineered microbes are designed to execute targeted functions, such as in situ drug synthesis, immune modulation, and targeted pathogen suppression within the GI milieu [5].
The advent of CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) and its associated Cas proteins has been transformative for microbial genome engineering, offering unparalleled precision, efficiency, and programmability. CRISPR-based tools enable the highly controlled insertion, deletion, or regulation of genes in probiotic strains, thereby facilitating the creation of smart probiotics capable of sensing environmental cues and producing therapeutic molecules in situ [6]. For instance, the commensal strain Escherichia coli Nissle 1917 (EcN) has been successfully modified to express anti-inflammatory agents, tumor-targeting proteins, and metabolic regulators, demonstrating efficacy across preclinical models of cancer, IBD, and metabolic disorders [7, 8].
The utility of CRISPR systems extends well beyond DNA editing. Emerging platforms, such as CRISPR-Cas13 and Cas7-11, permit RNA-level modulation, offering a crucial path for transient and reversible control over gene expression without permanent genomic alteration [9]. Furthermore, tools like CRISPR-associated transposases (CAST) and Cascade-Cas3 systems facilitate the insertion and deletion of large DNA segments, significantly expanding the scope of probiotic engineering to include complex synthetic circuits and multi-gene payloads [10]. These innovations have been vital for constructing advanced safety features, including microbial biosensors, self-destruct mechanisms, and quorum-sensing modules, which are essential for enhancing the safety and controllability of therapeutic applications [11].
CRISPR-engineered smart probiotics can be precisely customized to synthesize various therapeutic agents, such as short-chain fatty acids (SCFAs), anti-cancer peptides, and immunomodulatory cytokines, often in direct response to disease-specific biomarkers. Illustratively, Lactobacillus plantarum has been engineered to produce indole-3-lactic acid, significantly boosting the efficacy of immune checkpoint inhibitors in cancer models [12]. Similarly, modified Bacillus subtilis strains have been shown to increase butyrate production, exerting anti-obesity and anti-inflammatory effects in murine studies [13]. These examples underscore the immense potential of CRISPR-based probiotics to function as living drug factories, capable of delivering exquisitely precise treatments directly at the site of pathology.

FIGURE 2 Bacteria in disease diagnosis and therapy.
This figure illustrates the diverse roles of bacteria in disease diagnosis and treatment. Probiotics, genetically engineered bacteria, and the native microbiota contribute significantly to healthcare. Engineered bacteria can produce therapeutic proteins or metabolites, act as tumour vaccines, and activate immune responses, providing innovative cancer treatments. In the gastrointestinal tract, bacteria secrete beneficial compounds such as short- chain fatty acids, restoring healthy microbiota and treating intestinal diseases. Furthermore, bacteria can alleviate conditions such as periodontitis and diabetes by modulating the human microbiota. Additionally, bacteria function as biosensors, detecting diseases through various signals. These signals can be captured using optoacoustic imaging (OA), positron emission tomography (PET), magnetic resonance imaging (MRI), and wireless transmission or by analysing faecal samples.
The successful clinical translation of these applications relies heavily on efficient delivery of the CRISPR components into gut-resident microbes. Two principal strategies are currently being explored: the ex vivo engineering of probiotic strains followed by oral administration, and the in situ editing of native microbial populations directly within the GI tract. Engineered chassis strains like EcN and Lactobacillus rhamnosus have been successfully modified using CRISPR/Cas9 and CRISPR/Cas10 to suppress antibiotic resistance genes, modulate the tumor microenvironment, and generate therapeutic metabolites [15]. Concurrently, bacteriophage-based systems are being developed for the targeted delivery of CRISPR constructs into the gut, enabling the selective elimination of pathogenic strains and resistance plasmids while preserving commensal communities [15].
Despite these remarkable scientific strides, several challenges persist. Safety, stability, and regulatory compliance of genetically modified probiotics are paramount for clinical advancement. As noted by Leeflang et al., the inherent evolutionary adaptability of gut bacteria—manifesting as hypermutation, horizontal gene transfer, and clonal interference—can lead to unintended genetic changes, potentially jeopardizing therapeutic safety or efficacy [16]. Researchers are actively exploring strategies such as directed evolution, synthetic kill-switch circuits, and inducible promoters to maximize control and minimize off-target effects.
Moreover, the regulatory classification of next-generation probiotics remains inconsistent across jurisdictions. In the United States, CRISPR-engineered probiotics are typically regulated as LBPs under the FDA’s drug framework, mandating rigorous preclinical and clinical evaluation. Conversely, in Europe, the absence of harmonized guidelines results in non-uniform regulatory pathways, which complicates both product development and market entry [15]. Closing these regulatory gaps is critical for translating CRISPR-based microbial therapeutics into safe and effective clinical interventions.
This review synthesizes the current scientific understanding of CRISPR-based engineering of probiotics for targeted in situ drug synthesis. It explores the molecular mechanisms of diverse CRISPR systems, the sophisticated design principles of smart probiotics, the various delivery strategies, and the burgeoning therapeutic applications across metabolic, inflammatory, and neoplastic diseases. By integrating key insights from synthetic biology, microbiome science, and therapeutic design, CRISPR-based smart probiotics constitute a transformative frontier in precision medicine, poised to fundamentally redefine drug delivery and disease management from within the host.
Table 1- Comparison between next-generation probiotics and traditional probiotics.
|
|
NEXT GENERATION PROBIOTICS |
TRADITIONAL PROBIOTICS |
|
ORIGIN |
Derived from next-generation microorganisms that have been recently isolated using advanced tools and techniques |
Long history of use and are derived from a limited number of species, such as Lactobacillus and Bifidobacterium |
|
DEVELOPMENT |
Developed based on comparative analysis of microbiota compositions between healthy and diseased individuals |
Developed through a top-down screening strategy, which involves screening microbes enriched in healthy individuals compared to those in diseased individuals |
|
SAFETY |
Their safety is not yet proven as they are relatively new and have not been used for as long as traditional probiotics |
A long history of safe use in humans
|
|
APPLICATION |
Primarily used to treat or cure disease conditions |
Mainly used as food ingredients or supplements. |
|
REGULATION |
Considered to be live biotherapeutic products (LBPs) or drugs, which are subject to pharmaceutical clinical trials and research on their pharmacokinetics and pharmacodynamics |
They are not subject to the same level of regulation |
|
STRAIN SPECIFICITY |
Their health-promoting features are more closely tied to specific strains rather than entire species |
They are species-specific |
Overview of CRISPR-Cas9 Delivery Systems and Safety Considerations
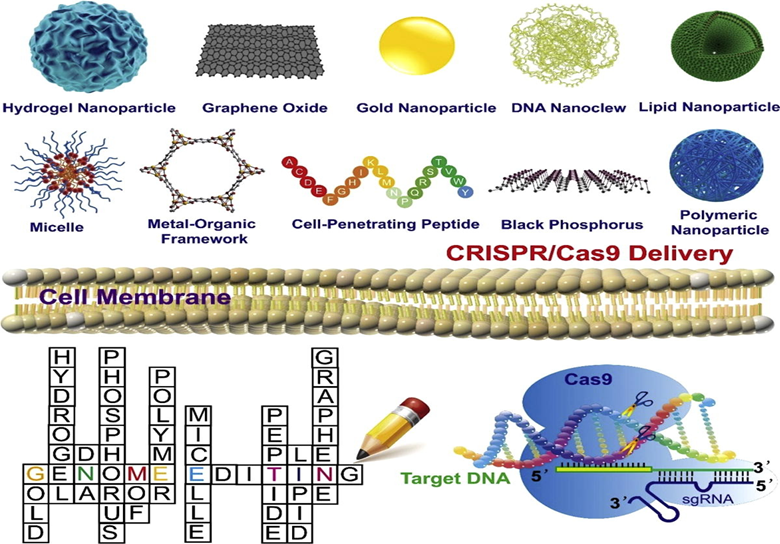
Figure-3 Overview of CRISPR-Cas9 Delivery Systems
The CRISPR-Cas9 system has revolutionized genome editing by enabling precise, programmable modifications across diverse life forms. Its clinical and agricultural potential, however, is inextricably linked to the development of robust and safe delivery strategies. The Cas9 system can be introduced in three main formats: plasmid DNA encoding Cas9 and single-guide RNA (sgRNA), messenger RNA (mRNA) encoding Cas9 with sgRNA, and pre-assembled Cas9-sgRNA ribonucleoprotein (RNP) complexes. Each format presents a distinct balance of editing kinetics, immunogenicity, and risk of off-target activity [17].
Plasmid-based delivery is favored for its simplicity and stability, often allowing co-expression of the entire machinery and repair templates. However, the large size of plasmids (>7 kb) is a barrier to efficient cellular and nuclear uptake. Critically, sustained Cas9 expression from plasmids can increase both off-target activity and systemic immune responses [17, 18]. mRNA delivery facilitates a rapid onset of editing due to immediate cytoplasmic translation and offers transient Cas9 expression, which generally reduces off-target effects. Yet, mRNA's inherent instability and susceptibility to degradation necessitate chemical modifications to enhance its stability and translational efficiency in vivo [17, 19]. RNP delivery provides the most immediate editing activity and the minimal off-target effects due to its purely transient nature. The main challenges here are the large size and charge of the Cas9 protein hindering cellular uptake, and the necessity for scrupulous management of endotoxin contamination during protein purification [17, 20].
Delivery vectors are broadly classified into physical, viral, and non-viral systems. Physical methods—such as microinjection, electroporation, and hydrodynamic tail vein injection—are highly effective in vitro or in small preclinical models but are inherently impractical for large-scale clinical application due to scalability and safety issues [17, 21]. Viral vectors, including adeno-associated virus (AAV), adenovirus, and lentivirus, offer high transduction efficiency and tissue tropism. Their use is, however, restricted by packaging capacity limitations, the potential for insertional mutagenesis, and immunogenicity. For example, AAV vectors have a restrictive maximum capacity of approximately 4.7 kb, often necessitating the use of smaller Cas9 orthologs like SaCas9 or complex split-system approaches [18, 22].
Non-viral vectors—such as lipid nanoparticles (LNPs), polymers, cell-penetrating peptides (CPPs), and gold nanoparticles—are increasingly attracting attention due to their modularity, enhanced safety profile, and potential for targeted delivery. LNPs are particularly promising for mRNA and RNP delivery, providing vital protection against enzymatic degradation and actively facilitating endosomal escape. Further modifications, such as PEGylation and ligand conjugation, can extend their circulation time and improve tissue specificity [17, 23]. Polymers like PEI and PLGA have been used for plasmid and mRNA delivery, though ongoing concerns surrounding biocompatibility and toxicity must be addressed. CPPs enable direct membrane translocation but may be susceptible to degradation or interference with sgRNA binding. Gold nanoparticles offer stability and tunable surface chemistry, and have successfully demonstrated in vivo RNP delivery with verified homology-directed repair (HDR) activity [17, 24].
Safety remains the core concern in CRISPR-Cas9 delivery. Off-target editing can result from prolonged Cas9 expression or non-specific sgRNA binding. The delivery format critically influences bioavailability and specificity; transient formats like RNPs are generally considered safer than persistent plasmid or viral systems [17, 25]. Furthermore, targeting specificity is dependent on the vehicle's ability to precisely localize the payload to the intended tissue. While direct injection has been employed for localized editing (e.g., in muscle), broader therapeutic strategies require advanced passive or ligand-directed delivery systems [17, 26].
Beyond delivery, safety considerations extend to the use of probiotics as the CRISPR chassis. As highlighted by Merenstein et al., the safety of LBPs requires meticulous assessment of strain identity, genome integrity, and the potential for antibiotic resistance gene transfer. Whole genome sequencing is essential for the identification of potential virulence factors and mobile genetic elements. The risk of horizontal gene transfer (HGT), especially involving clinically critical antibiotic resistance genes, must be rigorously evaluated using both genotypic and phenotypic assays. Regulatory bodies, including EFSA and FDA, strongly advise excluding strains that possess transferable resistance to key clinical antibiotics [27].
While rare, invasive infections linked to probiotic use have been documented, primarily in immunocompromised patients or those with indwelling medical devices. Although most cases respond well to standard antimicrobial therapy, the true incidence is likely underestimated due to underreporting and the difficulty of strain-level identification in clinical settings [27]. While not yet standard, microbiome profiling may eventually aid in predicting individual safety outcomes and therapeutic responses.
Recent innovations continue to expand the CRISPR toolkit. Zhong et al. introduced LrCas9, a Cas9 ortholog derived from the probiotic Lactobacillus rhamnosus GG, which recognizes a 5'-NGAAA-3' PAM sequence. LrCas9 has demonstrated superior editing efficiency in several important crop species compared to SpCas9-NG, SpRY, and LbCas12a, supporting multiplexed editing, base editing, and transcriptional regulation. Its reduced temperature sensitivity enhances its robustness for plant genome engineering, while its probiotic origin suggests potential safety advantages for future therapeutic applications [28].
In conclusion, the successful translation of CRISPR-Cas9 technology in both clinical and agricultural contexts hinges on the convergence of highly efficient delivery, precise biological targeting, and the most rigorous safety evaluation. Continuous advancements in non-viral vectors, probiotic strain engineering, and the discovery of novel Cas orthologs are vital for expanding the scope and reliability of genome editing technologies.
CRISPR-Cas9 Delivery Systems
The efficient delivery of CRISPR-Cas9 components is an absolute necessity for successful genome editing, especially in therapeutic applications. The chosen system must effectively bypass multiple biological barriers, ensure adequate cellular uptake, and simultaneously minimize adverse off-target effects. CRISPR-Cas9 can be delivered in three principal formats: plasmid DNA, mRNA, and ribonucleoprotein (RNP) complexes, each presenting distinct advantages and limitations [29].
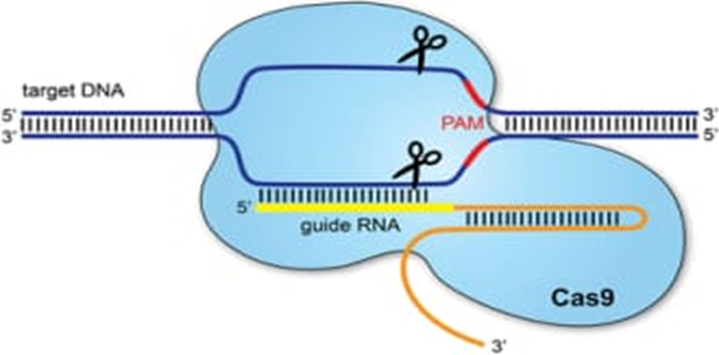
Figure 4- The CRISPR/Cas9 system.
Clustered regularly interspaced palindromic repeats (CRISPR) refers to sequences in the bacterial genome. They afford protection against invading viruses, when combined with a series of CRISPR-associated (Cas) proteins. Cas9, one of the associated proteins, is an endonuclease that cuts both strands of DNA. Cas9 is directed to its target by a section of RNA. This can be synthesised as a single strand called a synthetic single guide RNA (sgRNA); the section of RNA which binds to the genomic DNA is 18–20 nucleotides. In order to cut, a specific sequence of DNA of between 2 and 5 nucleotides (the exact sequence depends upon the bacteria which produces the Cas9) must lie at the 3’ end of the guide RNA: this is called the protospacer adjacent motif (PAM). Repair after the DNA cut may occur via two pathways: non-homologous end joining, typically leading to a random insertion/deletion of DNA, or homology directed repair where a homologous piece of DNA is used as a repair template. It is the latter which allows precise genome editing: the homologous section of DNA with the required sequence change may be delivered with the Cas9 nuclease and sgRNA, theoretically allowing changes as precise as a single base-pair.
1. Delivery Formats
Plasmid DNA (pDNA): This format encodes the Cas9 enzyme and the sgRNA in a single stable construct, offering cost-effectiveness. The primary drawback is its substantial size (>7 kb), which impedes cellular uptake and nuclear transport. Prolonged Cas9 expression from pDNA significantly elevates the risk of off-target activity and immunogenicity [29].
mRNA and sgRNA: This bypasses the need for nuclear transcription, enabling the rapid onset of editing activity. The transient Cas9 expression effectively reduces off-target activity, although the inherent instability of mRNA and its short expression duration can limit overall editing efficiency. Chemical modifications are used to enhance stability [29].
RNP Complexes: The pre-assembled Cas9 protein and sgRNA offer the fastest editing kinetics and the lowest demonstrated off-target activity. However, practical challenges include the large size of the protein, high production costs, and potential immunogenicity stemming from the bacterial origin of the Cas9 enzyme [29].
2. Delivery Technologies
Delivery methods are broadly categorized into physical, viral, and non-viral systems.
2.1 Physical Methods
Microinjection: Components are directly introduced into the nucleus or cytoplasm using fine needles. While offering high precision, it is labor-intensive and not feasible for large-scale in vivo or clinical applications [29].
Electroporation: Electric pulses are used to transiently permeabilize the cell membrane, allowing the entry of nucleic acids or proteins. Although effective in vitro and in zygotes, the high voltage can induce significant cell damage, which severely restricts clinical utility [29].
Hydrodynamic Tail-Vein Injection (HTVI): This method involves the rapid delivery of a large solution volume into the bloodstream, transiently increasing cell permeability. While effective in rodent liver, kidney, and lung tissues, it is not currently considered safe for human use [29].
2.2 Viral Vectors
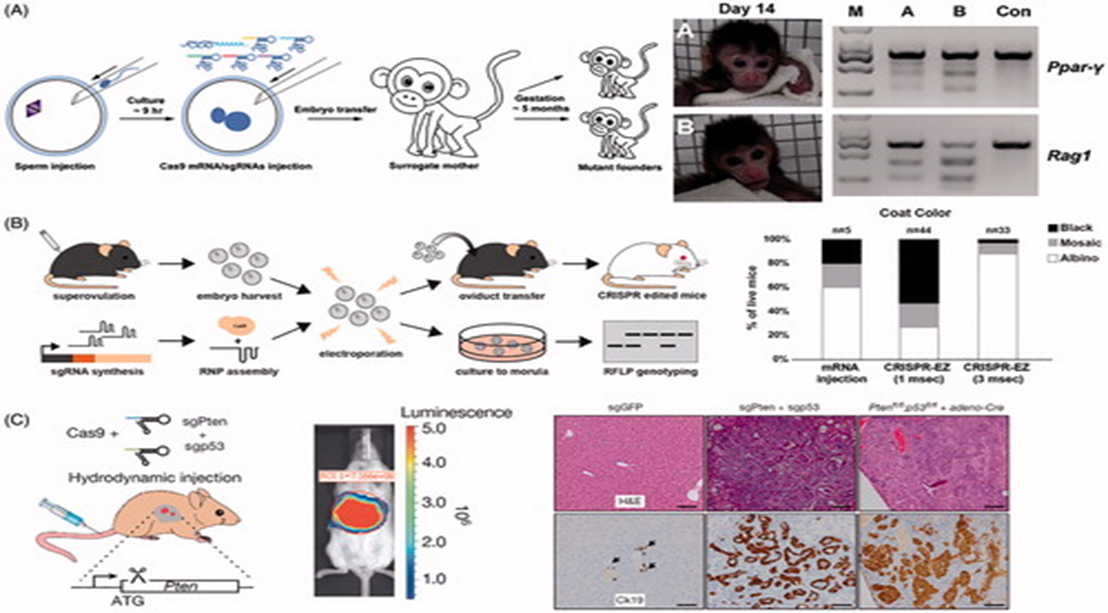
figure -5. Viral vector methods for delivery of CRISPR.
(A) AAV delivery of Cas9 and sgRNAs disrupting mutations in the Dmd gene in adult mdx mice, resulting in improvement of muscle biochemistry and function. Adapted with permission from Long et al. (2016). Copyright 2016 American Association for the Advancement of Science. (B) AAV intratracheal instillation delivery of sgRNAs in Cre-dependent Cas9 knock-in mice, resulting in lung adenocarcinoma (EGFP-positive tumors). Adapted with permission from Platt et al. (2016). Copyright 2014 Elsevier Inc. (C) A split Cas9 system in which the Cas9 C-terminal is packaged into one AAV vector and the Cas9 N-terminal is packaged into a second AAV vector. Reconstitution results in a fully functioning Cas9. Reprinted from Truong et al. (2015). Copyright 2014 The Authors (CC BY license). (D) AdV delivery of Cas9 and sgRNA targeting the Pten gene in mouse liver resulting in Pten mutation (see arrows by gel), and massive hepatomegaly and features of NASH in infected livers. Immu
Lentivirus: Characterized by high cloning capacity and efficient delivery of Cas9 and sgRNA. Major risks include insertional mutagenesis and potential oncogenicity. It has been used successfully to silence hepatitis B virus genes in hepatoma cells [29].
Adenovirus: A non-integrating vector with a natural tropism for the liver. It has been used to edit the PTEN gene in mouse liver, but concerns remain regarding long-term expression leading to possible hepatomegaly [29].
Adeno-Associated Virus (AAV): Currently the most widely used viral vector, offering targeted integration, low immunogenicity, and long-term expression in non-dividing cells. The main limitation is its small packaging capacity (\sim 4.7 kb), often necessitating the use of dual-vector systems or compact Cas9 variants like SaCas9 [29].
2.3 Non-Viral Vectors
Non-viral systems are generally considered safer and more flexible, although they often present with lower overall efficiency.
Polymers: Cationic polymers, such as PEI and PBAE, form protective complexes with nucleic acids. Functionalized polymers are being developed to improve targeting and reduce toxicity. Nanocapsules and micelles show promise in retinal and tumor therapies [29].
Lipid Nanoparticles (LNPs): These are efficient carriers for DNA, mRNA, and RNPs. LNPs can be surface-modified (e.g., with PEG or targeting ligands) to improve targeted delivery and stability, and to enhance lysosomal escape. Novel LNPs, such as X-O14B, have been designed specifically to improve RNP delivery [29].
Cell-Penetrating Peptides (CPPs): Facilitate the safe and targeted delivery of CRISPR components. However, they are prone to degradation and may interfere with Cas9-sgRNA binding due to complex charge interactions [29].
DNA Nano-Clew: Utilizes DNA nanowires to deliver Cas9-sgRNA complexes. While effective in vitro, further research is needed to assess long-term cytotoxicity and immunogenicity [29].
Gold Nanoparticles (AuNPs): Enable intracellular delivery via Au-S bonds. Hypoxia-responsive AuNPs have been successfully used for tumor-specific CRISPR release. In situ polymerization strategies are also being explored to improve RNP stability [29].
3. Delivery Challenges and Specificity
Key hurdles include achieving robust tissue-specific delivery, mitigating potential immune responses, and ensuring the minimal occurrence of off-target effects. The chosen delivery format is critical for editing specificity, with transient RNP systems generally showing reduced off-target risks compared to persistent viral or plasmid systems. Targeted delivery remains a significant technical challenge, with most current systems relying on either passive or ligand-based targeting strategies [30].
4. Microbiome-Driven Considerations
The design of therapeutic interventions is increasingly integrating knowledge of the gut microbiome, which significantly influences drug metabolism, immune responses, and disease progression. Personalized microbiome profiling holds the potential to guide future CRISPR-based interventions, especially for GI and metabolic disorders. However, the substantial inter-individual variability in microbiome composition remains a complex factor affecting consistent therapeutic outcomes [31].
CRISPR-Based Probiotics: Engineering the Next Generation of Microbial Therapeutics

Figure-6. Health benefits of next-generation probiotics
The development of microbial therapeutics is undergoing a profound transformation, evolving from the conventional use of probiotics—live microorganisms providing a health benefit when consumed in adequate amounts [34, 33]—to the creation of genetically engineered Next-Generation Probiotics (NGPs) [32]. Historically, probiotics have been utilized for managing gastrointestinal disorders such as IBD and antibiotic-associated diarrhea [34]. Nonetheless, conventional strains are often limited by poor stability within the harsh GI environment, a lack of precise targeting, and an inability to tightly regulate therapeutic agent delivery [34]. This convergence of challenges has catalyzed the application of synthetic biology, with the CRISPR-Cas system emerging as the most potent technology for constructing sophisticated, programmable microbial agents. CRISPR-based probiotics represent a critical subset of NGPs, leveraging genetic precision to transform beneficial bacteria into smart diagnostic and therapeutic vehicles [36, 38].
Definition and Concept of CRISPR-Based Probiotics
CRISPR-based probiotics are live microorganisms, typically strains of commensal or probiotic bacteria, that have been genetically modified using CRISPR and its associated Cas systems to perform specific, targeted therapeutic functions in situ [38]. The foundational concept involves harnessing the exceptional efficiency, precision, and specificity of the CRISPR-Cas system to either edit the native bacterial genome or introduce novel synthetic gene circuits [35]. The engineering aims to substantially augment the bacteria’s inherent capabilities, improving their stability, functionality, and specificity, thus enabling highly targeted and controlled therapeutic delivery [34]. Unlike simple drug carriers, these organisms are designed to be "smart," capable of sensing specific pathological biomarkers or environmental shifts within the host to modulate their behavior and therapeutic output [34]. This capacity for targeted gene modulation allows these engineered microbes to address complex medical challenges, ranging from combating multidrug-resistant pathogens to treating chronic conditions like cancer and metabolic disorders [36].
Molecular Mechanisms of CRISPR in Probiotics
The power of CRISPR-based probiotics is rooted in the molecular mechanism of the CRISPR-Cas system, an inherently specific and efficient genome editing tool [35]. The mechanism relies on a guide RNA (gRNA) that directs the Cas nuclease enzyme to a complementary target DNA sequence, which then induces a specific cleavage or modification [37]. This molecular precision allows researchers to implement highly specific edits within the probiotic genome [35].
CRISPR-Cas systems are structurally diverse, classified into two major classes and six further types [35]. Class 2 systems, particularly Type II (Cas9) and Type V (Cas12/Cpf1), are preferred for engineering due to their reliance on a single effector protein, which simplifies their introduction into host cells [35]. In the context of probiotic engineering, the technology is employed in several distinct ways:
Genome Modification: CRISPR-Cas can be used to permanently modify the probiotic host’s genetic material, facilitating the knock-in of novel synthetic circuits (e.g., for producing therapeutic molecules) or the targeted deletion of undesired genes [35].
Targeted Pathogen Elimination: The system's high specificity can be programmed to recognize and disrupt genes essential for the survival of target pathogenic bacteria, such as those encoding antibiotic resistance [36].
Pathway Reprogramming: The technology enables the modification of microbial metabolic pathways, allowing the engineered strain to specifically synthesize and secrete desirable therapeutic molecules, including proteins, peptides, or metabolites [32].
CRISPR-Cas systems are naturally present in various probiotic strains, including those in the Lactobacillus and Bacteroides fragilis genera [35]. Engineering efforts leverage these native systems or introduce well-characterized heterologous systems, such as Cas9 from Streptococcus pyogenes (SpCas9), to achieve the desired gene manipulation [35, 37].
Design Principles of Smart Probiotics
The creation of a functional CRISPR-based probiotic relies heavily on sophisticated synthetic biology principles to produce truly programmable microorganisms [38]. The objective of this engineering is to develop autonomous therapeutic systems that overcome the fundamental lack of precise delivery and control characteristic of conventional probiotic use [34].
Inducible Sense-and-Respond Systems
A crucial design element is the integration of synthetic gene circuits that enable the engineered probiotic to sense and respond to specific in situ cues [34]. These inducible systems ensure that the therapeutic action—be it gene editing or drug production—is triggered only when necessary, thereby maximizing efficacy and minimizing potential off-target effects [38]. These regulatory systems can be programmed to respond to various biological and environmental triggers, including:
Safety and Control Mechanisms
Given that CRISPR-based probiotics are live, genetically modified organisms, safety is paramount. The design must incorporate robust control mechanisms to mitigate potential risks, particularly the possibility of horizontal gene transfer (HGT) of the engineered constructs to non-target bacteria and the prevention of uncontrolled, off-target editing [34, 35]. A key strategy involves the use of drug-inducible CRISPR/Cas systems [35]. By utilizing ligand-binding domains from nuclear receptors, for instance, the activity of a split Cas enzyme can be tightly regulated by the administration of an external chemical signal [35]. Furthermore, synthetic biology facilitates the development of fail-safe mechanisms, such as self-destruct or "kill-switch" circuits, which ensure that the engineered bacteria are eliminated from the host upon completion of their therapeutic task or if they proliferate outside the designated target area [34, 38].
Therapeutic Applications of Engineered Probiotics
The precision afforded by CRISPR-Cas engineering has expanded the therapeutic utility of probiotics across a broad spectrum of diseases, enabling highly targeted interventions in areas previously inaccessible to conventional microbial therapy.
Metabolic Diseases
Engineered probiotics present a promising new avenue for managing complex metabolic disorders, such as diabetes and obesity. The microbes can be reprogrammed to function as in situ micro-factories that synthesize and deliver regulatory molecules [32]. For example, studies have demonstrated the capacity to incorporate therapeutically modified bacteria into the gut microbiota to actively inhibit obesity [32]. More ambitiously, modified commensal bacteria have been successfully leveraged to reprogram host intestinal cells, enabling them to become glucose-responsive, insulin-secreting cells for the targeted treatment of diabetes [32]. This cell-reprogramming capability highlights the potential of engineered microbes to move beyond simple drug delivery to fundamentally alter host physiological responses [36].
Inflammatory Diseases
Inflammatory conditions, including IBD, are traditional targets for probiotic intervention [34]. CRISPR-based engineering enhances this function by enabling the direct, local delivery of high-potency anti-inflammatory agents to the affected mucosal surface [32]. Specific strains, such as Streptococcus gordonii, have been engineered to achieve in vivo mucosal delivery of bioactive human Interleukin-1 Receptor Antagonist (IL-1ra), a powerful anti-inflammatory cytokine [32]. Beyond the GI tract, similar approaches have shown success in non-GI mucosal inflammation, such as using Lactococcus lactis strains engineered to secrete human Trefoil Factor 1 (TFF1) as a mouth rinse for the safe and effective treatment of oral mucositis [32].
Neoplastic Diseases (Cancer)
The intricate relationship between the human microbiome and cancer management is an intensive area of research, with the microbiome influencing metabolic pathways and immune functions that directly affect tumor progression [36]. Engineered probiotics are being developed to exploit this connection by performing critical functions within the tumor microenvironment:
Delivery Strategies for CRISPR Components
The clinical success of CRISPR-based probiotics critically depends on the development of efficient and targeted strategies for delivering the Cas/gRNA machinery to the target site, whether for ex vivo engineering or true in situ editing [37]. Current research is focused on three principal strategies:
This is the most established approach, involving the modification of the probiotic host in a laboratory setting [35]. CRISPR-Cas systems are used to achieve stable integration of the therapeutic synthetic circuit or desired gene edit into the bacteria’s genome [35], creating the "smart probiotic" chassis. After engineering and rigorous validation, the strains are cultivated, purified, and formulated for oral administration [34]. This process requires robust delivery vehicles to ensure the survival of the live cells through the extremely acidic stomach and the bile-rich small intestine [34]. To address this instability, significant effort is dedicated to creating advanced polymeric carriers and biopolymer encapsulation techniques to protect the engineered microbes until they reach the target location in the lower GI tract [34].
A technically demanding yet highly desirable approach is in situ editing, which eliminates the need for large-scale ex vivo cultivation and delivery of an externally engineered strain [38]. This strategy involves delivering the CRISPR machinery directly into the host's existing, native gut microbes to edit their genomes inside the body [38]. While this method offers the ultimate precision in modulating the entire microbial community, it faces significant challenges related to the efficient delivery of large Cas complexes and gRNAs across the complex bacterial cell wall and membrane in vivo [37]. Developing highly effective and selective delivery systems that can successfully operate within the dense, heterogeneous microbial environment remains a major obstacle for this precision medicine strategy [38].
Bacteriophages (phages) offer a natural, highly specific mechanism for introducing genetic material into bacterial cells [37]. Phage-based delivery exploits these bacteria-specific viruses as highly targeted delivery vehicles for CRISPR-Cas components [38]. The phage is engineered to carry the necessary Cas genes and gRNA, then administered to the host. Upon encountering the target bacteria, the phage injects the payload, triggering the desired editing event. This approach holds immense promise for the highly targeted editing and selective elimination of specific undesirable bacteria, such as pathogens, within the complex native microbiota [38]. Phage-mediated delivery possesses a distinct advantage due to its high natural infectivity and specificity, which can overcome many of the cellular barriers faced by non-viral delivery methods [37].
Safety, Control, and Regulatory Considerations
The emergence of CRISPR-based probiotics introduces complex safety and ethical considerations that mandate the development of rigorous regulatory frameworks [34, 32]. The primary safety concerns revolve around the potential for horizontal gene transfer (HGT) of the engineered genetic payload to other non-target microbes, including pathobionts like Bacteroides fragilis, which naturally possess CRISPR-Cas systems [35]. Furthermore, ensuring the long-term stability of the engineered strain and the precise control of its therapeutic output are critical [34].
To manage these risks, the integration of layered control mechanisms is essential [34]. The use of drug-inducible systems, where the Cas or therapeutic gene expression is tightly controlled by an external chemical signal or ligand [35], provides the clinician with a crucial safety switch [34]. For example, utilizing nuclear receptor ligand-binding domains facilitates the tight regulation of split CRISPR activity, ensuring that editing is confined to a defined therapeutic window [35].
From a regulatory standpoint, the development of NGPs utilizing synthetic biology tools introduces complexities that go beyond those of conventional probiotics [34]. Regulatory agencies must establish new, comprehensive safety assessment frameworks to evaluate the long-term ecological impact of these genetically modified organisms on the gut microbiome [34]. Crucially, the potential for HGT and the off-target effects of the CRISPR system must be thoroughly scrutinized before these powerful therapeutic tools can be safely and widely adopted in clinical practice [32, 34].
Design Principles of CRISPR-Engineered Smart Probiotics

Figure -7Primary mechanisms of action of probiotics against infections
(1) Probiotics produce antimicrobial substances like bacteriocins that cause cell death by inhibition of pathogen cell wall synthesis, (2) can enhance the barrier properties of epithelium enhancement of epithelial barrier by an interaction between MAMPs (i.e., LPS, CPS, and LTA) on the surfaces of probiotics and pattern recognition proteins on the epithelial barrier or modulation of intercellular junctions such as TJs, AJs, and desmosomes, and (3) can modulate the immune responses by interacting with dendritic cells. LPS: lipopolysaccharide; CPS: cell-wall-associated polysaccharide; LTA: lipoteichoic acid; MAMPs: microorganism-associated molecular patterns; TLRs: toll-like receptors; CLRs: C-type lectin receptors; TJs: tight junctions; AJs: adherence junctions. 59 Copyright ©2023 The authors. Published by the American Chemical Society. Reprinted (Adapted) with permission from Nilufer Yuksel, Busra Gelmez, and Ayca Yildiz-Pekoz. Molecular Pharmaceutics 2023, 20(7), 3320−3337 DOI: 10.1021/acs.molpharmaceut.3c00323.
Synthetic Circuitry and CRISPR Integration
The sophisticated design of CRISPR-engineered smart probiotics is predicated upon modular genetic architectures that enable environmental sensing, therapeutic expression, and controlled release. CRISPR-Cas systems—specifically Cas9, Cas12a, and their catalytically inactive variants (dCas9)—have been cleverly repurposed to regulate gene expression, perform genome editing, and facilitate base conversions in prominent probiotic chassis like Escherichia coli Nissle 1917 (EcN), Lactobacillus spp., and Bifidobacterium spp. [38, 39, 44].
Gene circuits are typically built using inducible promoters that respond to specific gut signals, such as nitric oxide (NO), tetrathionate, thiosulfate, or quorum-sensing molecules. These promoters ensure that the expression of CRISPR components or therapeutic payloads is activated only under disease-associated conditions. For instance, EcN strains have been engineered to secrete microcins or nanobodies upon sensing tetrathionate, a metabolite known to be elevated during inflammation [38]. Similarly, thiosulfate-responsive promoters have been utilized to regulate the secretion of immunomodulatory proteins in IBD models [39].
CRISPR interference (CRISPRi) and activation (CRISPRa) systems, which utilize dCas9 fused to transcriptional repressors or activators, allow for the precise modulation of gene expression without inducing DNA double-strand breaks (DSBs). These non-cleaving systems are especially valuable for finely controlling biosynthetic pathways and therapeutic gene expression within the probiotic chassis [39, 44].
Plasmid Stability and Containment Strategies
The issues of plasmid loss and genetic instability remain significant challenges in probiotic engineering. To counteract this, strategies such as the chromosomal integration of CRISPR components and the implementation of toxin-antitoxin stabilization systems have been employed [38, 41]. Additionally, the creation of auxotrophic strains and the use of kill-switch circuits—such as passive suicide modules or inducible lysis systems—have been developed as essential means to prevent uncontrolled proliferation and subsequent horizontal gene transfer [44, 45].
Containment strategies also focus on using plasmids devoid of traditional antibiotic resistance markers, favoring alternative selection systems such as the upp gene or Cre/loxP recombination with lox72 sites, which help to avoid genetic scars and reduce recombination risks [39, 52].
Off-Target Control and Editing Precision
Off-target activity remains a critical safety concern in all CRISPR applications. To address this, high-fidelity Cas9 variants and optimized guide RNA scaffolds have been developed to significantly enhance targeting specificity [40, 54]. Base editors, derived from Cas9 or Cas12a, enable precise single-nucleotide conversions without inducing DSBs, thereby reducing overall genomic disruption and improving the safety profile of the engineered strains [40, 54].
Mechanism of Action
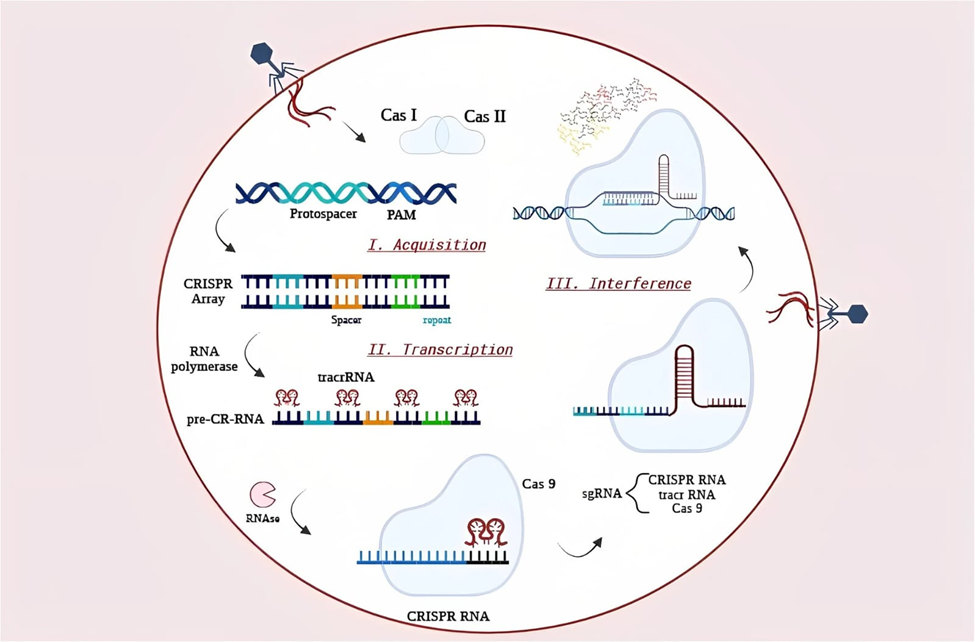
Figure-8 Mechanism of CRISPR
CRISPR-engineered probiotics function through a carefully orchestrated, multi-step cascade involving sensing, activation, expression, and therapeutic release. Upon successfully colonizing the gastrointestinal tract, the engineered strains detect disease-associated signals via their synthetic biosensors. These signals activate the inducible promoters linked to the CRISPR modules, thereby initiating a precise transcriptional or post-transcriptional response [38, 39].
For instance, EcN strains engineered with NO-responsive promoters secrete type III interferons to actively suppress inflammation in IBD models [39]. Similarly, tetrathionate-sensing circuits trigger the release of microcins designed to target and eliminate pathogenic Enterobacteriaceae [38].
CRISPR-Cas9 systems induce DSBs at target loci, which are then repaired via non-homologous end joining (NHEJ) or homology-directed repair (HDR), enabling targeted gene knockouts or precise edits [38, 58]. Conversely, dCas9-based systems modulate transcription without cleavage, while base editors (e.g., cytosine or adenine deaminases fused to Cas9) enable targeted nucleotide conversions [40, 54].
Therapeutic payloads—such as antimicrobial peptides, cytokines, or metabolic enzymes—are delivered via native transporters, engineered secretion systems, or programmed lysis. Outer membrane vesicles (OMVs) and bacterial ghosts are also being utilized as delivery platforms, capable of encapsulating drugs or immunomodulators for highly targeted release [45, 64].
Therapeutic Applications
CRISPR-modified probiotics have demonstrated substantial preclinical efficacy in treating IBD, ulcerative colitis, and Crohn’s disease. EcN strains engineered to secrete trefoil factors, anti-TNF$\alpha$ nanobodies, and SCFA biosynthetic enzymes have shown notable mucosal healing and inflammation suppression in murine colitis models [38, 39].
Metabolic disorders, including phenylketonuria, hyperammonemia, and hyperoxaluria, have been addressed using engineered EcN strains capable of degrading toxic metabolites. In preclinical studies, CRISPR-edited Bacillus subtilis strains producing butyrate were shown to significantly improve insulin sensitivity and lipid profiles in obese mice [47].
Engineered probiotics are being deployed to combat multidrug-resistant pathogens through the secretion of bacteriocins and microcins. EcN strains expressing microcin I47 or J25 have successfully eradicated ESBL-producing E. coli and carbapenem-resistant Klebsiella pneumoniae in vivo [38, 43].
In oncology, CRISPR-modified EcN strains expressing Tum5 or anti-CD47 nanobodies have demonstrated tumor regression and enhanced immune infiltration in murine models [44, 49]. OMVs functionalized with melanin and calcium phosphate shells have also enabled effective photothermal therapy with reduced systemic toxicity and improved survival in tumor-bearing mice [45].
Probiotic strains have been engineered to express viral antigens or immunomodulators for respiratory infections. Lactobacillus paracasei engineered to express phospholipase D reduced lung inflammation and ACE2 expression in SARS-CoV-2 challenged mice [63]. Furthermore, OMVs displaying influenza peptides have been shown to confer protection against H1N1 and H3N2 in murine models [65].
Challenges and Ethical Considerations
Technical challenges persist, including plasmid instability, metabolic burden, and off-target effects. Strategies such as chromosomal integration, toxin-antitoxin systems, and the use of high-fidelity Cas variants are continuously being developed to address these issues [38, 40].
Biosafety concerns primarily revolve around horizontal gene transfer, unintended colonization, and potential immunogenicity. Robust containment strategies—suching auxotrophy, kill switches, and non-antibiotic selection markers—are essential to mitigate these risks [39, 44, 52].
Ethical debates center on the manipulation of commensal microbes and the long-term ecological impact of releasing engineered strains into the human body and the environment. Regulatory ambiguity regarding classification and approval pathways further complicates clinical translation [53].
Public perception and acceptance of microbial genome editing remain critical, necessitating transparent communication, comprehensive public education, and the provision of robust safety data. Personalized approaches, guided by individual microbiome profiling, may help address the significant interindividual variability in therapeutic response.
FUTURE PERSPECTIVES
Next-generation CRISPR tools, such as prime editing and sophisticated CRISPRa/i systems, offer expanded capabilities for precise, programmable modulation of probiotic genomes without inducing DSBs [54]. These systems enable a level of dynamic control over gene expression and therapeutic output previously unattainable.
The integration with AI and multi-omics platforms promises to significantly refine strain selection, predict host responses, and optimize complex circuit design. Machine learning models have already been applied to classify disease states based on microbiome profiles and guide targeted therapeutic development [15].
Regulatory evolution is an essential prerequisite for supporting clinical translation. Harmonized international guidelines, standardized safety assessments, and clear approval pathways are urgently needed. Collaborative efforts among researchers, clinicians, and regulatory bodies will be paramount for thoughtfully navigating the ethical and ecological dimensions of this transformative field.
In summary, CRISPR-engineered smart probiotics represent a profoundly transformative approach to targeted drug delivery within the gut. Continued technological innovation, rigorous scientific validation, and thoughtful, responsive regulation will ultimately determine their trajectory from cutting-edge experimental tools to indispensable clinical therapeutics.
REFERENCE

Shraddha Shinde, Pooja Rasal, Pranav Sonawane, Pranav Taskar, Shubham Adhav, CRISPR-Engineered Smart Probiotics: A Novel Approach to Targeted Gut Therapeutics, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 11, 2979-2999. https://doi.org/10.5281/zenodo.17651329
 10.5281/zenodo.17651329
10.5281/zenodo.17651329
