1,3,4,5 Division of Parasitology, ICMR - National Institute for Research in Bacterial Infections (ICMR-NIRBI), Kolkata, India.
2 Department of Biomedical Chemistry, School of International Health, Graduate School of Medicine, The University of Tokyo, Tokyo, Japan.
Andrographolide (ADG), a bioactive compound found in Andrographis plants, exhibits significant antigiardial effects by down-regulating gene expression at both transcriptomic and proteomic levels. Giardiasis, caused by Giardia lamblia, is a common intestinal infection, particularly in developing countries. We investigated the impact of ADG on Giardia pathogenesis using in vitro co-culture models with human intestinal epithelial cells (INT-407). ADG did not show any cytotoxic effect on normal intestinal cells. However, in Giardia-infected co-cultures, ADG treatment effectively eliminated trophozoites and restored epithelial structural integrity. Real-time PCR analysis revealed that ADG significantly downregulated seven virulence-related genes, including those involved in oxidative stress, cell structure, metabolism, and immune evasion. The cathepsin B, a secreted cysteine protease implicated in host tissue damage and immune modulation, showed reduced expression at both mRNA and protein levels, particularly after 6 to 9 hours of ADG exposure. These findings indicate that ADG disrupts Giardia’s survival mechanisms by impairing key virulence pathways. The dual-level gene expression modulation by ADG suggests its strong potential as a natural therapeutic agent against giardiasis, particularly for drug-resistant strains, and provides new insight into the molecular mechanisms of Giardia pathogenesis and drug response.
Giardiasis, caused by the protozoan parasites Giardia lamblia, which primarily colonizes the upper small intestine, is a globally prevalent intestinal infection affecting both humans and animals (Gardner et al. 2001). Despite its widespread prevalence, the mechanisms and mediators of pathogenesis, including the molecular characterization of strain virulence, remain largely unclear. Most Giardia infections remain asymptomatic, though the parasite primarily colonizes the duodenum, causing localized villus atrophy and epithelial apoptosis at infection sites. An estimated 280-million giardiasis cases have been diagnosed worldwide every year, and the prevalence rates of the infection may vary between 20-30% in developing countries and 2-5% in developed countries, respectively (Haldar et al. 2024). Giardiasis presents variably in humans, from asymptomatic cases to acute symptoms including diarrhoea, abdominal cramps, nausea, and vomiting. Giardiasis mainly affects the duodenum but also harms the wider gut, leading to diarrhoea not limited to the infection site (Cotton et al. 2014). Cathepsin B, a cysteine protease with a unique occluding loop, may damage the duodenum by acting as endo- or exopeptidase (Turk et al. 2012). Giardia has 27 cathepsin protease genes, but most of their functions are unknown. Some may help the parasite evade the immune system. A recent study revealed that Giardia trophozoites secrete cathepsin B-like proteases, which may help the parasite evade the immune system by degrading intestinal IL-8 and reducing the host's inflammatory response (Cotton et al. 2014). Giardia’s secreted cathepsin B protease may break down intestinal mucin and help trophozoites attach to the gut lining.
Andrographolide (ADG) is a principal bioactive compound of the Andrographis plant and has been used in conventional treatment for various diseases. ADG demonstrates an extremely broad spectrum of biological activities. Recent reports suggest that it possesses anti-tumour, cardioprotective, antioxidant, anti-inflammatory, cytotoxic, immunomodulatory, antibacterial, neuroprotective, anti-HIV, and hepatoprotective properties (Haldar et al. 2024). Furthermore, it has been shown to exhibit antimicrobial activity against bacteria and viruses. In addition to its documented properties, Andrographolide has been identified for its effectiveness against Leishmania (Sinha et al. 2000), Plasmodiaum (Mishra et al. 2011), dengue (Edwin et al. 2016), and protozoan infections (Banerjee et al. 2017).
Our study showed ADG’s effects on the expression of various genes involved in Giardia pathogenesis, such as oxidative stress, cell cycle, metabolic pathways, and other regulatory genes, assessed at both the RNA and protein levels.
Methodology
Parasite culture
Giardia trophozoites were cultured in a modified TYI-S-33 medium, which had been filter-sterilized and supplemented with 10% adult bovine serum (Raj et al. 2014). The cultures were maintained at 37°C under microaerophilic conditions and sub-cultured upon reaching confluence. To harvest the parasites, the culture medium was removed and replaced with fresh medium for routine culture. Subcultures were maintained every 48 hours and only trophozoites from the logarithmic phase were used for all experiments.
Mammalian cell line culture (INT-407)
Human intestinal epithelial (INT-407) cells were cultured in minimal essential medium containing Earle's salts (Gibco) at 37°C in a 5% CO2 atmosphere (Sousa et al. 2001). The growth medium, composed of α-Minimum Essential Medium, was supplemented with 10% fetal bovine serum (FBS) and antibiotics (10 units/ml penicillin and 10 μg/ml streptomycin). To sustain cell viability, the media were renewed every 2 days. Upon achieving confluency, cell harvesting was conducted using trypsin-EDTA (Sigma). The resulting confluent monolayers were then utilized for various applications, including electrophysiological experiments, co-culture experiments with Giardia parasites.
INT-407 and Giardia Co-culture
Confluent monolayers of INT-407 cells were utilized in the experiment. The cell media in the INT-407 monolayers was removed and replaced with a mixture comprising 90% complete DMEM and 10% Giardia medium, with or without the addition of Giardia trophozoites (105 total parasites). To prevent any potential contamination by parasites, control cultures were carefully maintained in a separate plate. Microscopic examination was conducted on the control flask to confirm the absence of Giardia cross-contamination. Subsequently, the co-cultures were placed in an incubator at 37°C with 5% CO2 for a duration of 24 hours. Following this incubation period, the Giardia parasites were eliminated (Dubourg et al. 2018). In a brief procedure, the culture media was withdrawn from the flask and the INT-407 cell media was substituted with a blend of 99.9% complete DMEM and 0.1% Giardia medium, with or without the inclusion of Giardia supernatant.
RNA Extraction
RNA was extracted from ~2×10? trophozoites in the log phase using 1 ml Trizol. After pelleting and 5-minute incubation, 0.2 ml chloroform was added per 1 ml Trizol, followed by 15 seconds of mixing and 5-minute incubation. Centrifugation (12,000g, 20 minutes, 2-8°C) separated the phases. The aqueous phase was transferred and RNA was precipitated with 0.5 ml isopropanol and a 10-minute incubation (15-30°C). After centrifugation (12,000g, 10 minutes, 2-8°C), the RNA pellet was washed with 75% ethanol, centrifuged (7,500g, 5 minutes, 2-8°C), air-dried, and dissolved in 20 μl DEPC water.
cDNA Preparation
cDNA synthesis was performed using the Bio Bharati muLV Kit per manufacturer’s instructions in a 20 µl reaction using 10 ng total RNA. The mix included 1 µl RNA, 1 µl oligo dT, 1 µl dNTP mix, and 9 µl water (total 12 µl), incubated at 65°C for 5 minutes and cooled on ice. Then, 4 µl first-strand buffer, 1 µl RNase inhibitor, and 2 µl 0.1M DTT were added and incubated at room temperature. Next, 1 µl Super RT (200 U/µl) was added, and the reaction incubated at 42–52°C for 50 minutes, followed by inactivation at 70°C for 5 minutes.
Real Time PCR
Numerous virulence factors play a crucial role in Giardia pathogenesis. Seven specific genes-Arginine deiminase, NADH, Kinase coding gene, FtsJ, Cathepsin, Cysteine-rich membrane protein, and Surface protein, are selected for transcriptomic analysis to confirm their differential expression using real-time PCR. These genes were chosen for their roles in biochemical and molecular activities. Expression levels are assessed by RT-qPCR using a beta actin-related housekeeping gene for normalization. PCR began with incubation at 95°C for 15 minutes, followed by 40 cycles of 94°C for 15 seconds (denaturation), 60°C for 30 seconds (annealing), and 72°C for 30 seconds (extension) for DNA amplification.
Western blot
Western blot analysis was used to evaluate the effect of cathepsin B gene on Giardia trophozoites during activation. After SDS-PAGE, proteins were transferred to a nitrocellulose membrane at 100 V for 1 hour. Membranes were blocked with 5% skim milk in TBST for 1 hour at room temperature. Primary antibody (Cathepsin B, 1:1000) was incubated overnight at 4?°C. After three TBST washes, membranes were incubated with anti-mouse secondary antibody (1:20,000) for 2 hours at room temperature. Protein bands were visualized using enhanced Fastcast™ Acrylamide solution (Bio Bharati).
RESULTS
Co-culture study
The study demonstrated that the ADG did not induce any significant changes in normal INT-407 intestinal tissue culture when administered alone. This suggests that, under the tested conditions, ADG does not exert noticeable effects on normal intestinal cells. However, in a distinct scenario involving a co-infection with both intestinal cells and Giardia trophozoites, a notable impact was observed (Figure 1). The Giardia trophozoites caused disruption to the epithelial junction between cells in this co-infection. After treatment with ADG, co-infection for 24 hours resulted in the complete removal of Giardia trophozoites, accompanied by observable changes in the cell line. This suggests that ADG possesses potent activity against Giardia trophozoites, leading to their elimination and subsequent alterations in the intestinal cell line.
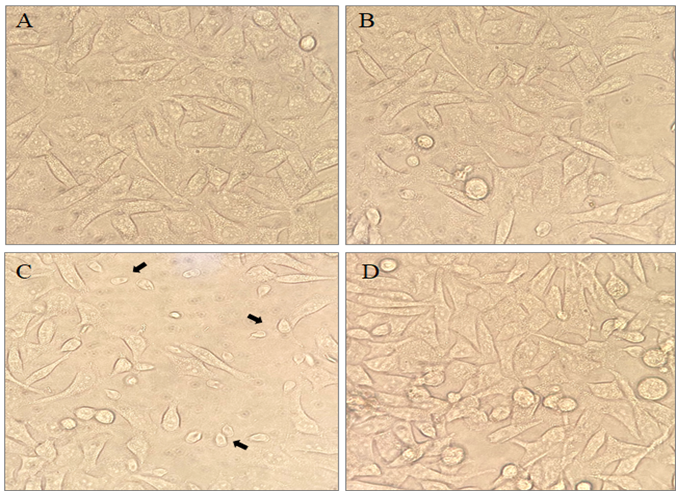
Figure 1: The effects of ADG on Co-culture: (A) Control INT-407 intestinal cells. (B) INT-407 cells treated with ADG. (C) Co-infection of Giardia with INT-407 cells, and (D) ADG treatment on co-infected cells.
Gene expression changes
We examined the expression of seven genes by RT-qPCR to assess mRNA fold changes after ADG treatment. As shown in Figure 2, gene expression varied after 12 hours of treatment with ADG (5?μM concentration). Overall, ADG treatment consistently downregulated the expression of all seven genes linked to Giardia pathogenicity.
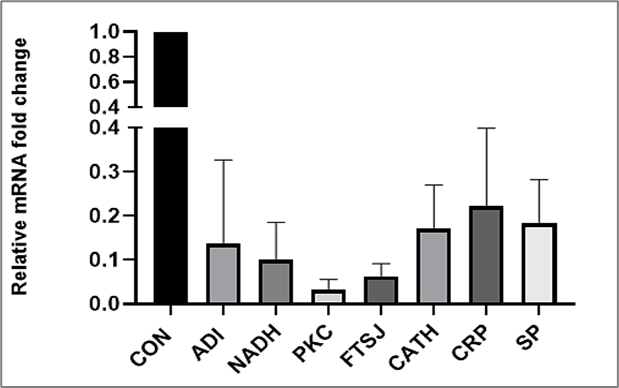
Figure 2: qRT-PCR analysis of mRNA fold changes in Giardia trophozoites treated with ADG.
Cathepsin B Expression
ADG-induced changes in cathepsin B expression vary across different time intervals. Our results show that gene expression depends on the duration of treatment. After 3 hours of ADG exposure, no significant changes were observed; however, substantial expression changes were detected between 6 and 9 hours (Figure 3). These findings reveal that ADG significantly alters cathepsin B expression at the proteomic level over various time points.
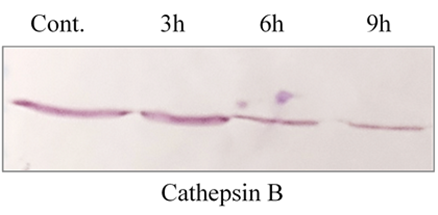
Figure 3: The expression patterns of the cathepsin B gene were analyzed across various time intervals. Gene expression was assessed at specific time points- 3?h, 6?h, and 9?h at a particular concentration.
DISCUSSION
Andrographolide has been reported to have therapeutic effects for treating various diseases (Jayakumar et al. 2013). This plant contains a variety of chemical compounds, including diterpenoids, glycosides, flavonoids, lactones, and flavonoid glycosides. It has been reported that ADG possesses various biological activities, including antihepatitic, antidiarrheal, anti-inflammatory, anti-allergic, immunostimulatory, antidiabetic, antioxidant, and antimalarial properties (Hossain et al. 2014; Haldar et al 2024). Additionally, it has demonstrated hepatoprotective, anticancer, cardiovascular, antihyperglycemic (Hossain et al. 2014), and anti-HIV activities. The anti-inflammatory action of ADG involves the inhibition of nuclear transcription factor-(kappa) B, rendering it a potential therapeutic target for treating cancer and autoimmune diseases (Rajagopal et al. 2003). The previous study provided evidence supporting the primary mechanism of Giardia attachment through the ventral disk, with microtubules playing a significant role. This attachment was hindered by colchicine and mebendazole, both of which affect microtubular function (Sousa et al. 2001). We investigated treatment with ADG in both intestinal cells and co-culture, and observed inhibition of trophozoite attachment to intestinal cells without any adverse effects on the intestinal cells themselves.
In our previous study, we assessed the antigiardial activity of ADG against Giardia trophozoites through treatment with various doses and time intervals (Haldar et al. 2024). Our findings reveal that ADG substantially inhibited the expression of pathogenic factors associated with Giardia, inducing cell apoptosis like death and down regulating the expression of several genes. Additionally, ADG effectively inhibits the expression of pathogenic factor-regulating genes, including those involved in metabolism, cell division, surface protein regulation and cytoskeleton regulation. To our knowledge, this study is the first to identify ADG's ability to target various pathogenic factor-regulating genes during Giardia pathogenesis.
Recent studies have highlighted the significance of cathepsin B-like protease activities from Giardia in host-pathogen interactions (Beatty et al. 2017; Manko et al. 2017). However, the specific proteases involved have not been identified. Our study focuses on the expression of cathepsin-B genes, which belong to the Cysteine protease family, to identify proteomic-level expression changes following ADG treatment. Over different time intervals (3h, 6h and 9h), there was a gradual decrease in expression of cathepsin B.
CONCLUSION
Andrographolide inhibited the expression of various genes in Giardia trophozoites, particularly those involved in pathogenesis, at the transcriptomics and proteomics levels. Additionally, ADG suppressed the protein expression of the cathepsin B gene at different time point intervals. Furthermore, it inhibited the adherence of Giardia trophozoites in co-culture with INT-407 intestinal cells. These positive results help inform future drug development strategies and address global health challenges related to parasitic diseases.
REFERENCES



Tapas Haldar, Sanjib Kumar Sardar, Ajanta Ghosal, Arjun Ghosh, Sandipan Ganguly, Andrographolide Modulates Giardia Gene Expression at Transcriptomic and Proteomic Levels, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 5, 4978-4984. https://doi.org/10.5281/zenodo.15555070
 10.5281/zenodo.15555070
10.5281/zenodo.15555070
